Что такое фотосинтез и почему он так важен для нашей планеты
Фотосинтез — один из самых важных биологических процессов на Земле. Благодаря фотосинтезу живые организмы получают кислород, необходимый для дыхания, а сами растения создают полезные органические вещества для своей жизнедеятельности. В этой статье мы поговорим о том, что обозначает фотосинтез, как он происходит и что образуется в процессе фотосинтеза.

Время чтения:
Что такое фотосинтез
Фотосинтез — процесс, при котором в клетках, содержащих хлорофилл, под действием энергии света образуются органические вещества из неорганических. При фотосинтезе растение поглощает углекислый газ и воду, синтезирует органические вещества и выделяет кислород, как побочный продукт фотосинтеза.
Процессы фотосинтеза идут в тканях, содержащих хлоропласты, — преимущественно, в листе, на который приходится большая часть процессов фотосинтеза. Такая ткань называется хлоренхима, или мезофилл.
Строение хлоропластов
Чтобы понять, что происходит в растении при фотосинтезе, изучим подробнее хлоропласты. Хлоропласты — это особые пластиды растительных клеток, в которых происходит фотосинтез. Основные элементы структурной организации хлоропластов высших растений представлены на рис.1.

Хлоропласт — это двумембранный органоид. Внешняя мембрана проницаема для большинства органических и неорганических соединений. Она содержит специальные транспортные белки, благодаря которым нужные для работы хлоропласта пептиды и другие вещества попадают в него из цитоплазмы. Внутренняя мембрана обладает избирательной проницаемостью и способна контролировать, какие именно вещества попадут во внутреннее пространство хлоропласта.
Для хлоропластов характерна сложная система внутренних мембран, позволяющая пространственно организовать фотосинтетический аппарат, упорядочить и разделить реакции фотосинтеза, несовместимые между собой, и их продукты. Мембраны образуют тилакоиды, которые, в свою очередь, собираются в «стопки» — граны. Пространство внутри тилакоидов называется внутритилакоидным пространством, или люменом.
Внутреннее пространство хлоропласта между гранами заполняет строма — гидрофильный слабоструктурированный матрикс. В строме содержатся необходимые для реакций синтеза сахаров ферменты, а также рибосомы, кольцевая молекула ДНК, крахмальные зёрна.

Световая фаза фотосинтеза
Чтобы лучше понять, что происходит во время фотосинтеза, разберём фазы фотосинтеза. Световая фаза фотосинтеза включает в себя фотохимические и фотофизические процессы, и может быть поделена на три этапа:
- Фаза поглощения — энергия света улавливается при помощи светособирающих комплексов, переходит в энергию электронного возбуждения пигментов, передаётся в реакционный центр фотосистем I и II.
- Фаза реакционных центров — энергия электронного возбуждения пигментов светособирающих комплексов используется для активации реакционных центров фотосистем. В реакционном центре электрон от возбуждённого хлорофилла передаётся другим компонентам электрон-транспортной цепи, пигмент после отдачи электрона переходит в окисленное состояние и становится способным, в свою очередь, отнимать электроны у других веществ. Именно в этом процессе происходит преобразование физической формы энергии в химическую.
- Фаза электрон-транспортной цепи — электроны переносятся по цепи переносчиков, образуются АТФ, НАДФН, O2. Необходимо, чтобы каждый переносчик электрон-транспортной цепи поочерёдно восстанавливался и окислялся, обеспечивая таким образом перенос энергии электронов. Любой этап переноса электрона сопровождается высвобождением или поглощением энергии. Часть энергии теряется. На некоторых участках электрон-транспортной цепи перенос электрона сопряжён с переносом протона.
Для того чтобы понять, что происходит во время фазы фотосинтеза, рассмотрим эти процессы подробнее. Кванты света улавливаются светособирающими комплексами фотосистемы I — молекула хлорофилла в составе светособирающего комплекса переходит в возбуждённое состояние, и энергия передаётся в реакционный центр фотосистемы I. Происходит возбуждение молекул хлорофилла фотосистемы I, отщепляется электрон. Пройдя по цепочке внутренних компонентов фотосистемы I и внешних переносчиков, электрон в конце концов попадает к НАДФ+ — образуется восстановитель НАДФН. Получается, что хлорофилл фотосистемы I отдал электрон и приобрёл положительный заряд, и для дальнейшего функционирования необходимо восстановить нейтральность молекулы, получить электрон, чтобы закрыть «дырку». Этот электрон приходит от фотосистемы II.
На светособирающие комплексы фотосистемы II попадают кванты света — происходит возбуждение молекулы хлорофилла фотосистемы II, молекула хлорофилла отдаёт электрон и переходит в окисленное состояние. Нехватку электрона хлорофилл восполняет благодаря фотолизу воды, при этом образуется протоны H+, а также важный побочный продукт фотосинтеза — кислород. По цепи переносчиков электрон от хлорофилла фотосистемы II попадает к хлорофиллу реакционного центра фотосистемы I и восстанавливает его. Теперь этот хлорофилл может снова поглощать энергию кванта света и отдавать электрон в электрон-транспортную цепь.
Протоны, попадающие во внутритилакоидное пространство, используются для синтеза АТФ. С помощью фермента АТФ-синтазы за счёт градиента протонов образуется АТФ из АДФ и фосфата. Под градиентом понимают неравномерное распределение: во внутритилакоидном пространстве H+ больше, в строме — меньше. Поэтому частицы стремятся проникнуть в строму, переходят в неё через АТФ-синтазу, а в процессе пути сквозь белковый комплекс отдают ему часть энергии, которая и используется для синтеза АТФ.
Темновая фаза фотосинтеза
Что образуется при фотосинтезе в темновую фазу? В строме хлоропластов с помощью энергии АТФ и восстановителя НАДФН, полученных в световую фазу, образуются простые сахара, из которых в ходе других процессов образуется крахмал. Ферментативные процессы не нуждаются в наличии света. Важнейший процесс, происходящий в темновую фазу фотосинтеза, — фиксация углекислого газа воздуха. Синтез и превращения сахаров в хлоропластах имеют циклический характер и носят название цикл Кальвина.
В нём можно выделить три этапа:
- Фаза карбоксилирования (введение CO2 в цикл).
- Фаза восстановления (используются АТФ и НАДФН, полученные в световую фазу).
- Фаза регенерации (превращения сахаров).
В строме хлоропластов находится производное простого пятиуглеродного сахара рибозы. С помощью особого фермента (Рубиско) к производному рибозы присоединяется CO2 (реакция карбоксилирования) — образуется неустойчивое шестиуглеродное соединение, которое быстро распадается на две трехуглеродные молекулы. Дальше, с затратой АТФ и НАДФН, полученных в ходе световых процессов, трехуглеродное соединение модифицируется — образуется восстановленное соединение с атомом фосфора и альдегидной группой в составе. Теперь перед клеткой стоит проблема: необходимо получить шестиуглеродное соединение — глюкозу для синтеза крахмала, а также пятиуглеродное — производное рибозы для того, чтобы эти процессы могли начаться заново. Для решения этих проблем в фазу регенерации из полученных ранее трехуглеродных соединений под действием ферментов образуются четырёх-, пяти-, шести- и семиуглеродные сахара. Из шестиуглеродной молекулы образуется глюкоза, из которой синтезируется крахмал. Из пятиуглеродной молекулы образуется производное рибозы и цикл замыкается. Остальные сахара также используются клеткой в других биохимических процессах.
Отдельно стоит сказать про крайне важный фермент первой фазы цикла Кальвина — рибулозо-1,5-дифосфаткарбоксилазу (Рубиско). Это сложный фермент, состоящий из 16 субъединиц, с молекулярной массой в 8 раз больше, чем у гемоглобина. Является одним из важнейших ферментов в природе, поскольку играет центральную роль в основном механизме поступления неорганического углерода (из CO2) в биологический круговорот. Содержание Рубиско в листьях растений очень велико, он считается самым распространённым ферментом на Земле.

Значение фотосинтеза
В процессе фотосинтеза энергия света заключается в энергию химических связей органических веществ. Поэтому фотосинтез служит первичным источником почти всей энергии, используемой живыми организмами в процессе жизнедеятельности. Практически все живые организмы, за исключением хемосинтетиков, так или иначе пользуются теми продуктами, что выделяются при фотосинтезе.
За счёт фотосинтеза сформировалась и поддерживается пригодная для дыхания атмосфера с высоким содержанием кислорода.
Фиксация углекислого газа в ходе фотосинтеза служит главным местом входа неорганического углерода в биогеохимический цикл. Также ассимиляция CO2 препятствует перегреву Земли, предотвращая парниковый эффект.
Заключение
Каждый год на нашей планете благодаря фотосинтезу производится около 200 миллиардов тонн кислорода, из которого образуется озоновый слой, защищающий от ультрафиолетовой радиации. Фотосинтез помогает поддерживать состав атмосферы и препятствует увеличению количества углекислого газа. Без растений и кислорода, который они выделяют в процессе фотосинтеза, жизнь на нашей планете была бы просто невозможна.
Скоро перезвоним!
Или напишем на почту, если не получится дозвониться
Фотосинтез – что это, определение, как происходит, фазы, значение, фото и видео


Природа
Автор Вадим Хромов На чтение 14 мин Опубликовано 31.03.2020 Обновлено 02.12.2020
Леса считаются «зелеными легкими планеты» не напрасно. Благодаря постоянной работе растений, фотосинтезу, все живое на Земле имеет возможность дышать. Более того, без них простейшие организмы не смогли бы эволюционировать и сложно представить, какой облик имела бы планета. Что такое фотосинтез и как происходит данный процесс, рассмотрим в деталях.
Что такое фотосинтез?
Фотосинтез – биохимический процесс, во время которого с помощью особых пигментов растений и энергии света из неорганических веществ (углекислого газа, воды) возникают органические. Это один из наиболее важных процессов, за счет которого появилось и продолжает существовать большинство организмов на планете.

Интересный факт: к фотосинтезу способны наземные растения, а также зеленые водоросли. При этом водоросли (фитопланктон) вырабатывают 80% кислорода.
Значение фотосинтеза для жизни на Земле
Без фотосинтеза вместо множества живых организмов на нашей планете существовали бы одни лишь бактерии. Именно энергия, полученная в результате данного химического процесса, позволила бактериям эволюционировать.
Любые природные процессы нуждаются в энергии. Она поступает от Солнца. Но правильную форму солнечный свет приобретает лишь после того, как преобразовывается растениями.
Растения используют лишь часть энергии, а остальную накапливают в себе. Ими питаются травоядные животные, которые являются пищей для хищников. В ходе образовавшейся цепочки каждое звено получает необходимые ценные вещества и энергию.

Кислород, вырабатываемый в ходе реакции, необходим для дыхания всем существам. Дыхание представляет процесс, противоположный фотосинтезу. При этом органические вещества окисляются, разрушаются. Полученная энергия используется организмами для выполнения различных жизненно необходимых задач.
В период существования планеты, когда растений было мало, кислород практически отсутствовал. Примитивные формы жизни получали минимум энергии другими способами. Ее было слишком мало для развития. Поэтому дыхание за счет кислорода открыло более широкие возможности.
Еще одна функция фотосинтеза – защита организмов от воздействия ультрафиолетового света. Речь идет об озоновом слое, находящемся в зоне стратосферы на высоте около 20-25 км. Образуется он за счет кислорода, который превращается в озон под действием солнечного света. Без этой защиты жизнь на Земле ограничивалась бы только подводными организмами.
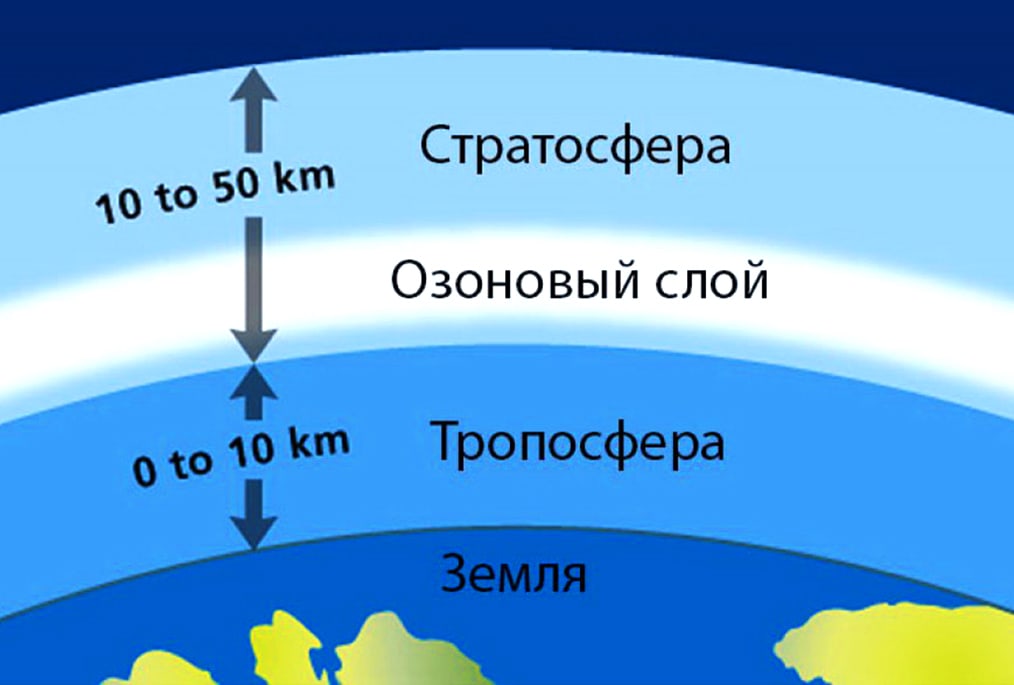
Организмы выделяют во время дыхания углекислый газ. Он является обязательным элементом фотосинтеза. В противном случае углекислый газ просто накапливался бы в верхних слоях атмосферы, значительно усиливая парниковый эффект.
Это серьезная экологическая проблема, суть которой состоит в повышении температуры атмосферы с негативными последствиями. К ним относится изменение климата (глобальное потепление), таяние ледников, повышение уровня Мирового океана и др.
- выделение кислорода;
- образование энергии;
- образование питательных веществ;
- создание озонового слоя.
Определение и формула фотосинтеза
Термин «фотосинтез» произошел от сочетания двух слов: фото и синтез. В переводе с древнегреческого они означают «свет» и «соединение» соответственно. Таким образом, энергия света превращается в энергию связей органических веществ.

Углекислый газ + вода + свет = углевод + кислород.
Научная формула фотосинтеза:
Фотосинтез происходит так, что непосредственный контакт воды и СО2 не наблюдается.
Значение фотосинтеза для растений
Растениям для роста и развития требуются органические вещества, энергия. Благодаря фотосинтезу они обеспечивают себя данными компонентами. Создание органических веществ – основная цель фотосинтеза для растений, а выделение кислорода считается побочной реакцией.

Интересный факт: растения уникальны, поскольку для получения энергии другие организмы им не нужны. Поэтому они образуют отдельную группу – автотрофы (в переводе с древнегреческого языка «питаюсь сам»).
Как происходит фотосинтез?
Фотосинтез протекает непосредственно в зеленых частях растений – хлоропластах. Они входят в состав растительных клеток. Хлоропласты содержат вещество – хлорофилл. Это и есть тот основной фотосинтетический пигмент, благодаря нему происходит вся реакция. Кроме того, хлорофилл определяет зеленый цвет растительности.
Интересно: Растения Красной книги России — описание, фото, видео, особенности, где растут
Для этого пигмента характерна способность поглощать свет. А в клетках растения запускается настоящая биохимическая «лаборатория», в которой вода и СО2 превращаются в кислород, углеводы.
Вода поступает через корневую систему растения, а газ проникает непосредственно в листья. Свет выступает в качестве источника энергии. Когда частица света действует на молекулу хлорофилла, происходит ее активация. В молекуле воды H2O кислород (O) остается невостребованным. Таким образом, он становится побочным для растений, но таким важным для нас, продуктом реакции.
Фазы фотосинтеза
Фотосинтез делится на две стадии: световую и темновую. Протекают они одновременно, но в разных частях хлоропласта. Название каждой фазы говорит само за себя. Световая или светозависимая фаза происходит только при участии частиц света. Темновой или светонезависимой фазе наличие света не требуется.
Прежде чем рассматривать каждую фазу подробнее, стоит разобраться в строении хлоропласта, поскольку оно определяет суть и место протекания стадий. Хлоропласт является разновидностью пластид и внутри клетки расположен отдельно от остальных ее компонентов. Он имеет форму зернышка.
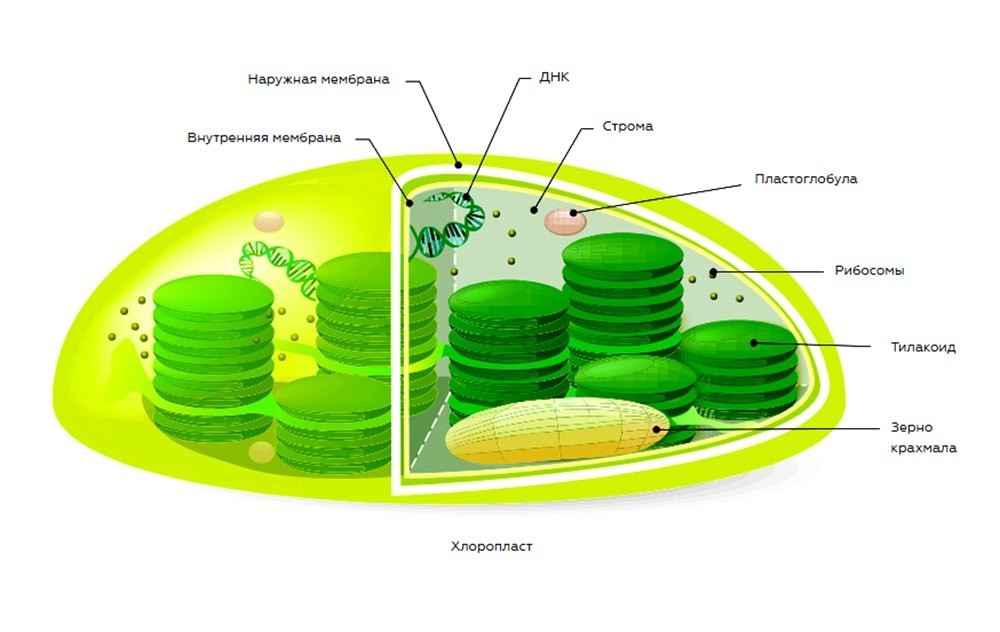
Составляющие части хлоропласта, участвующие в фотосинтезе:
- 2 мембраны;
- строма (внутренняя жидкость);
- тилакоиды;
- люмены (просветы внутри тилакоидов).
Световая фаза фотосинтеза
Протекает на тилакоидах, точнее, их мембранах. Когда на них попадает свет, выделяются и накапливаются негативно заряженные электроны. Таким образом, фотосинтетические пигменты лишаются всех электронов, после чего наступает очередь распада молекул воды:
При этом образованные протоны водорода имеют положительный заряд и копятся на внутренней мембране тилакоида. В итоге протоны с зарядом плюс и электроны с зарядом минус разделены лишь мембраной.
Происходит выработка кислорода, как побочного продукта:
В определенный момент фазы электронов и протонов водорода становится слишком много. Тогда в работу вступает фермент – АТФ-синтаза. Его задача состоит в том, чтобы переместить протоны водорода из мембраны тилакоида в жидкую среду хлоропласта – строму.

На этом этапе водород попадает в распоряжение другого переносчика – НАДФ (сокращение от никотинамиддинуклеотидфосфат). Это также разновидность фермента, который ускоряет окислительные реакции в клетках. В данном случае его работа состоит в транспортировке протонов водорода в реакции углеводов.
На данной стадии происходит процесс фотофосфолирования, во время него вырабатывается огромное количество энергии. Ее источником является АТФ – аденозинтрифосфорная кислота.
- Попадание кванта света на хлорофилл.
- Выделение электронов.
- Выделение кислорода.
- Образование НАДФН-оксидазы.
- Образование энергии АТФ.
Интересный факт: существует реликтовое растение под названием вельвичия, растущее на африканском побережье Атлантического океана. Это единственный представитель своего рода с минимумом листьев, способных к фотосинтезу. Однако возраст вельвичий достигает около 2000 лет.

Темновая фаза фотосинтеза
Светонезависимая фаза происходит непосредственно в строме. Она представляет собой ряд ферментативных реакций. Углекислый газ, поглощенный на световой стадии, растворился в воде, а на этом этапе он восстанавливается до глюкозы. Также вырабатываются сложные органические вещества.
Реакции темновой фазы делятся на три основных типа и зависят от вида растений (точнее, их метаболизма), в клетках которых происходит фотосинтез:
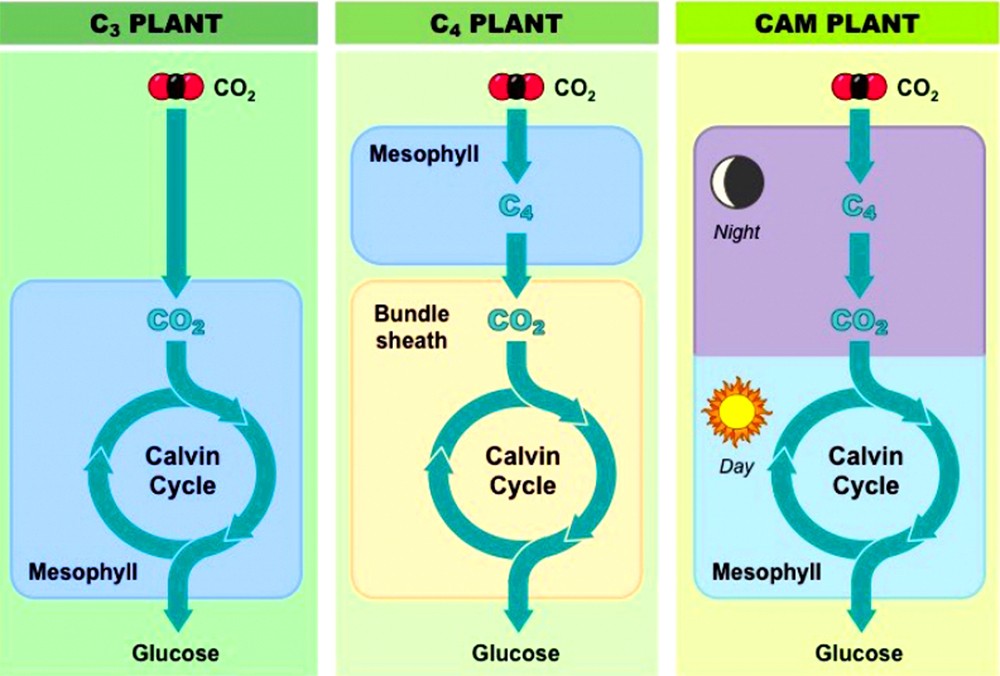
- С3-растения;
- С4-растения;
- САМ-растения.
К С3-растениям относится большая часть культур сельскохозяйственного назначения, которые растут в умеренном климате. В ходе фотосинтеза у них углекислый газ становится фосфоглицериновой кислотой.
К С 4 -растениям принадлежат субтропические и тропические виды, преимущественно сорняки. Для них характерна трансформация углекислого газа в оксалоацетат. САМ-растения – категория растений, которым не хватает влаги. Они отличаются особенным видом фотосинтеза – CАМ.
С3-фотосинтез
Наиболее распространенным является С3-фотосинтез, который также именуется циклом Кальвина – в честь американского ученого Мелвина Кальвина, который внес огромный вклад в изучение данных реакций и получил за это Нобелевскую премию.
Растения называются С3 из-за того, что во время реакций темновой фазы образуются 3-углеродные молекулы 3-фосфоглицериновой кислоты – 3-PGA. Непосредственное участие принимают различные ферменты.

Чтобы образовалась полноценная молекула глюкозы, должно пройти 6 циклов реакций светонезависимой фазы. Углевод – главный продукт фотосинтеза в цикле Кальвина, но помимо него вырабатываются жирные и аминокислоты, а также гликолипиды. У С3 растений фотосинтез проходит исключительно в клетках мезофилла.
Интересно: Климат: что такое, описание, виды, особенности, фото и видео
Главный недостаток С3-фотосинтеза
Растения, относящиеся к группе С3, характеризуются одним существенным недостатком. Если в окружающей среде отмечается недостаточный уровень влаги, способность к фотосинтезу существенно снижается. Это происходит по причине фотодыхания.
Дело в том, что при невысокой концентрации углекислого газа в хлоропластах (меньше 50:1 000 000) вместо фиксации углерода происходит фиксация кислорода. Специальные ферменты существенно замедляются и расходуют солнечную энергию впустую.
Одновременно с этим замедляется рост и развитие растения, поскольку оно недополучает органические вещества. Также не происходит выброс кислорода в атмосферу.
Интересный факт: морской слизень Elysia chlorotica – уникальное животное, которое осуществляет фотосинтез как растения. Оно питается водорослями, хлоропласты которых проникают в клетки пищеварительного тракта и фотосинтезируют там на протяжении месяцев. Вырабатываемые углеводы служат для слизня пищей.

С 4 -фотосинтез
В отличие от C3-синтеза, здесь реакции фиксации углекислого газа осуществляются в различных клетках растений. Эти виды растений способны справляться с проблемой фотодыхания, и делают они это при помощи двухэтапного цикла.
С одной стороны поддерживается высокий показатель углекислого газа, а с другой – контролируется низкий уровень кислорода в хлоропластах. Подобная тактика позволяет растениям С 4 избежать фотодыхания и связанных с ним сложностей. Представителями растений данной группы являются сахарный тростник, кукуруза, просо и др.
По сравнению с растениями С3 они способны намного интенсивнее выполнять процессы фотосинтеза при условии высокой температуры и недостатка влаги. На первом этапе углекислый газ фиксируется в клетках мезофилла, где образуется 4-углеродная кислота. Затем кислота переходит в оболочку и распадается там на 3-углеродное соединение и углекислый газ.

На втором этапе полученный углекислый газ начинает работать в цикле Кальвина, где вырабатывается глицеральдегид-3-фосфат и углеводы, необходимые для энергетического обмена.
Благодаря двухэтапному фотосинтезу в растениях С 4 образуется достаточное для цикла Кельвина количество углекислого газа. Поэтому ферменты работают в полную силу и не растрачивают энергию напрасно.
Но у и этой системы есть свои минусы. В частности расходуется больший объем энергии АТФ – она необходима для трансформации 4-углеродных кислот в 3-углеродные и в обратном направлении. Таким образом, С3-фотосинтез всегда продуктивнее, чем С 4 при должном количестве воды и света.
Что влияет на скорость фотосинтеза?
Фотосинтез может протекать с различной скоростью. Этот процесс зависит от условий окружающей среды:

- вода;
- длина волны света;
- углекислый газ;
- температура.
Вода является основополагающим фактором, поэтому при ее недостатке реакции замедляются. Для фотосинтеза наиболее благоприятны волны красного и сине-фиолетового спектра. Также предпочтительнее высокая степень освещенности, но лишь до определенного значения – при его достижении связь между освещенностью и скоростью реакции исчезает.
Высокая концентрация углекислого газа обеспечивает быстрые фотосинтетические процессы и наоборот. Определенная температура важна для ферментов, которые ускоряют реакции. Идеальные условия для них – около 25-30℃.
Фотодыхание
Дышать необходимо всем живым существам, и растения не являются исключением. Однако этот процесс у них происходит немного иначе, чем у людей и животных, отчего носит название фотодыхания.
В целом, дыхание – физический процесс, во время которого живой организм и окружающая его среда обмениваются газами. Как и всему живому, растениям для дыхания нужен кислород. Но потребляют они его гораздо меньше, чем вырабатывают.
В ходе фотосинтеза, который происходит только при солнечном свете, растения создают для себя пищу. Во время фотодыхания, которое осуществляется круглосуточно, эти питательные вещества ими поглощаются с целью поддержки метаболизма внутри клеток.
Интересный факт: в течение солнечного дня участок леса площадью 1 гектар потребляет от 120 до 280 кг углекислого газа и выделяет от 180 до 200 кг кислорода.
Кислород (как и углекислый газ) проникает в клетки растений через особые отверстия – устьица. Они располагаются в нижней части листочков. На одном листе может располагаться около 1000 устьиц.

Газообмен растений в зависимости от освещенности
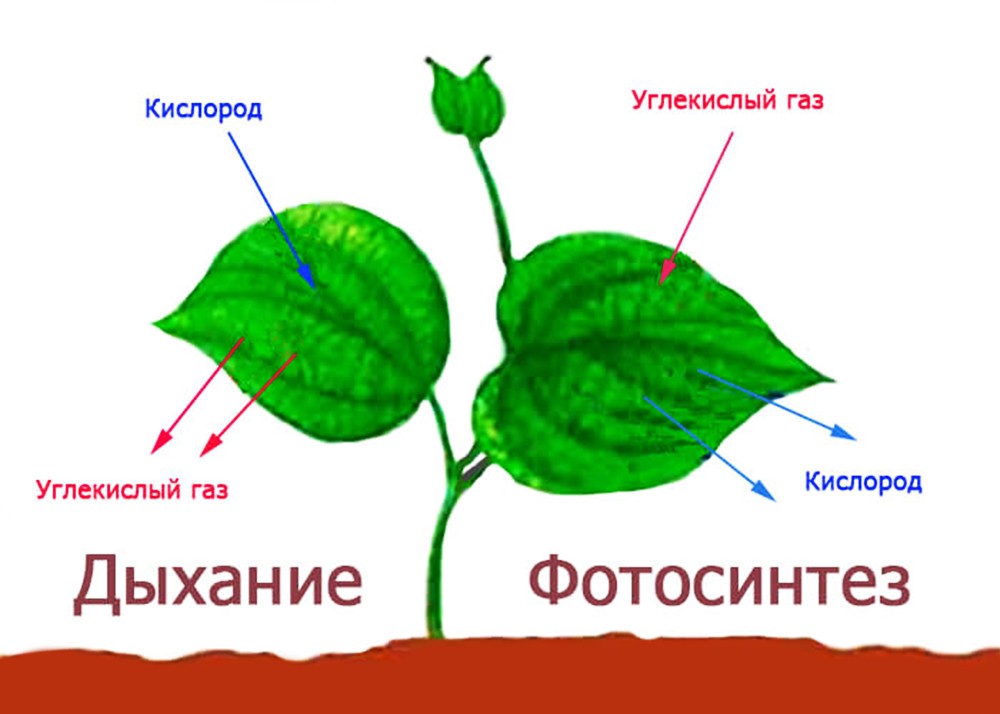
Процесс газообмена при разной освещенности представлен следующим образом:
- Яркий свет. Во время фотосинтеза используется углекислый газ. Растения выделяют больше кислорода, чем потребляют. Его излишки попадают в атмосферу. Углекислый газ потребляется быстрее, чем выделяется дыханием. Неиспользованные углеводы запасаются растением впрок.
- Тусклый свет. Газообмен с окружающей средой не происходит, поскольку растение потребляет весь кислород, который производит.
- Отсутствие света. Происходят только процессы дыхания. Углекислый газ выделяется, а кислород потребляется.
Интересно: Как измеряют скорость ветра?
Хемосинтез
Некоторые живые организмы тоже способны к образованию моноуглеводов из воды и углекислого газа, при этом они не нуждаются в солнечном свете. К ним относятся бактерии, а процесс преобразования энергии называется хемосинтезом.
Хемосинтез являет собой процесс, во время которого синтезируется глюкоза, но вместо солнечной энергии используются химические вещества. Протекает он в зонах с достаточно высокой температурой, подходящей для работы ферментов, и отсутствием света. Это могут быть области вблизи гидротермальных источников, утечек метана на морских глубинах и др.

Источником энергии для бактерий выступают химические связи метана и сероводорода. В результате хемосинтеза возникает сера и ее соединения в качестве побочных продуктов реакции.
История открытия фотосинтеза
История открытия и изучения фотосинтеза берет начало в 1600 г., когда Ян Батист ван Гельмонт решил разобраться в актуальном на тот момент вопросе: чем питаются растения и откуда они черпают полезные вещества?
В то время считалось, что источником ценных элементов является почва. Ученый поместил в емкость с землей веточку ивы, но предварительно измерил их вес. На протяжении 5 лет он ухаживал за деревом, поливая его, после чего снова провел измерительные процедуры.
Выяснилось, что вес земли снизился на 56 г, однако деревце стало в 30 раз тяжелее. Это открытие опровергло мнение о том, что растения питаются почвой и породило новую теорию – водного питания.

В дальнейшем многие ученые пытались ее опровергнуть. Например, Ломоносов считал, что частично структурные компоненты попадают к растениям через листья. Он руководствовался растениями, которые успешно растут на засушливых территориях. Однако доказать эту версию не удалось.
Ближе всего к реальному положению вещей оказался Джозеф Пристли – ученый-химик и священник по совместительству. Однажды он обнаружил погибшую мышь в перевернутой вверх дном банке, и этот случай заставил его провести в 1770-х годах ряд опытов с грызунами, свечами и емкостями.
Пристли обнаружил, что свеча всегда быстро тухнет, если накрыть ее сверху банкой. Также не может выжить и живой организм. Ученый пришел к выводу, что существуют некие силы, которые делают воздух пригодным для жизни, и попытался связать это явление с растениями.
Он продолжил ставить опыты, но в этот раз попробовал поместить под стеклянную емкость горшочек с растущей мятой. К огромному удивлению, растение продолжало активно развиваться. Тогда Пристли поместил под одну банку растение и мышь, а под вторую – только животное. Результат очевиден – под первой емкостью грызун остался невредим.

Достижение химика стало мотивацией для других ученых всего мира повторить эксперимент. Но загвоздка была в том, что священник проводил опыты в дневное время. А, к примеру, аптекарь Карл Шееле – ночью, когда появлялось свободное время. В итоге, ученый обвинил Пристли в обмане, ведь его подопытные не переносили эксперимент с растением.
Между химиками разразилось настоящее научное противостояние, которое принесло существенную пользу и дало возможность сделать еще одно открытие – чтобы растения восстанавливали воздух, им нужен солнечный свет.
Конечно, фотосинтезом это явление тогда еще никто не называл, да и оставалось немало вопросов. Однако в 1782 ботаник Жан Сенебье смог доказать, что при наличии солнечного света растения способны расщеплять углекислый газ на клеточном уровне. А в 1864, наконец, появилось экспериментальное доказательство того, что растения поглощают углекислый газ и выделяют кислород. Это заслуга ученого из Германии – Юлиуса Сакса.
Фотосинтез – интересное видео
Если Вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Что такое фотосинтез? Влияние углекислого газа CO2 на скорость фотосинтеза. Краткое пособие
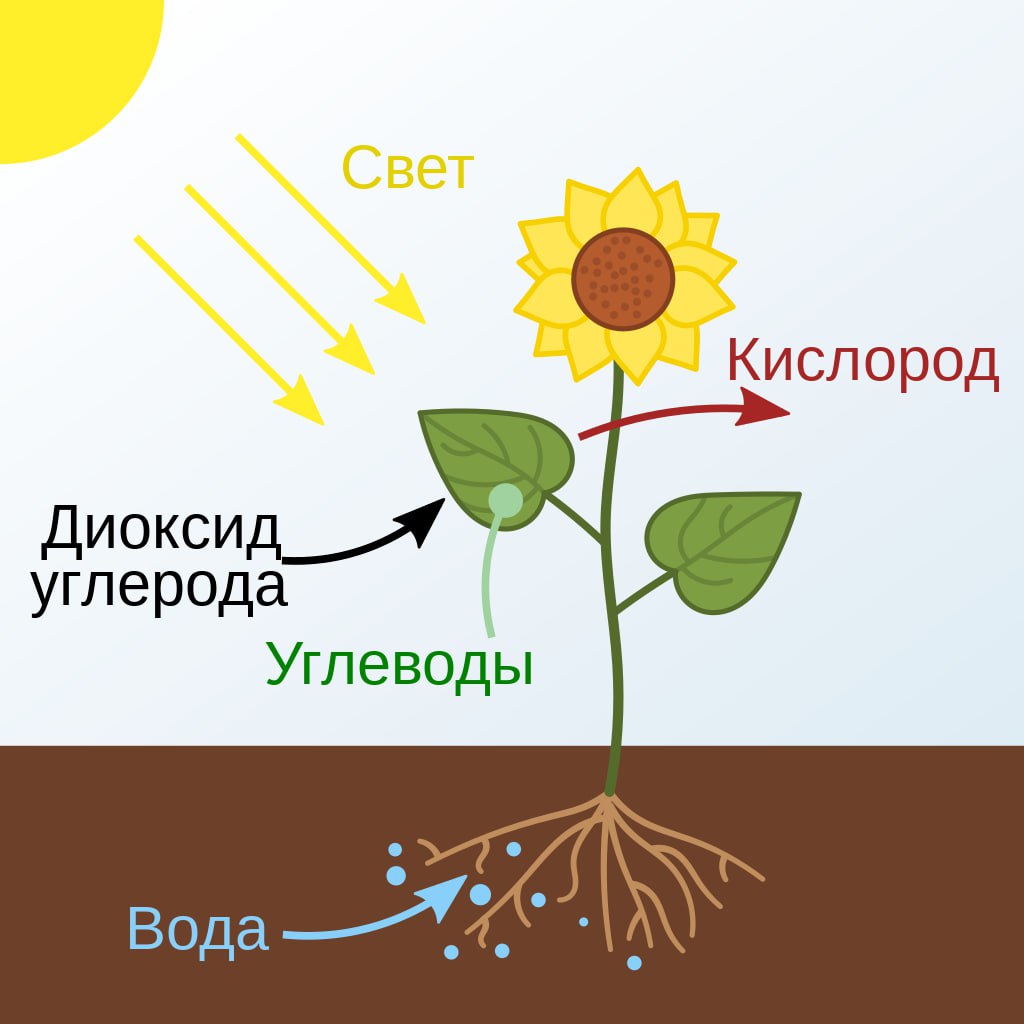
Фотосинтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов ( хлорофилл у растений , бактериохлорофилл у бактерий и бактериородопсин у архей ).
Фотосинтез — это процесс, благодаря которому существует большинство живых организмов на нашей планете. Фотосинтез — процесс образования органических веществ из углекислого газа (CO2) и воды (H2O), протекающий с использованием солнечной энергии.
Растение вырабатывает глюкозу и кислород. Для фотосинтеза растению необходимы вода, углекислый газ и свет: растение использует свет в качестве энергии для производства кислорода и глюкозы из воды и углекислого газа. Это основные строительные блоки, необходимые растению для роста.
Бесхлорофилльный фотосинтез
Система бесхлорофилльного фотосинтеза отличается значительной простотой организации, в связи с чем предполагается эволюционно первичным механизмом запасания энергии электромагнитного излучения. Эффективность бесхлорофилльного фотосинтеза как механизма преобразования энергии сравнительно низка (на один поглощённый квант переносится лишь один H + ).
Хлорофилльный фотосинтез
Хлорофилльный фотосинтез отличается от бактериородопсинового значительно большей эффективностью запасания энергии. На каждый эффективно поглощённый квант излучения против градиента переносится не менее одного H + , и в некоторых случаях энергия запасается в форме восстановленных соединений (ферредоксин, НАДФ).
Фотосинтез — процесс с крайне сложной пространственно-временной организацией.
Разброс характерных времен различных этапов фотосинтеза составляет 19 порядков: скорость процессов поглощения квантов света и миграции энергии измеряется в фемтосекундном интервале (10 −15 с), скорость электронного транспорта имеет характерные времена 10 −10 —10 −2 с, а процессы, связанные с ростом растений, измеряются днями (10 5 —10 7 с).
Также большой разброс размеров характерен для структур, обеспечивающих протекание фотосинтеза: от молекулярного уровня (10 −27 м 3 ) до уровня фитоценозов (10 5 м 3 ).
В фотосинтезе можно выделить отдельные этапы, различающиеся по природе и характерным скоростям процессов:
Лист
Фотосинтез растений осуществляется в хлоропластах — полуавтономных двухмембранных органеллах, относящихся к классу пластид.
Хлоропласты могут содержаться в клетках стеблей, плодов, чашелистиков, однако основным органом фотосинтеза является лист. Он анатомически приспособлен к поглощению энергии света и ассимиляции углекислоты. Плоская форма листа, обеспечивающая большое отношение поверхности к объёму, позволяет более полно использовать энергию солнечного света. Вода, необходимая для поддержания тургора и протекания фотосинтеза, доставляется к листьям из корневой системы по ксилеме — одной из проводящих тканей растения. Потеря воды в результате испарения через устьица и в меньшей степени через кутикулу (транспирация) служит движущей силой транспорта по сосудам. Однако избыточная транспирация является нежелательной, и у растений в ходе эволюции сформировались различные приспособления, направленные на снижение потерь воды.
Отток ассимилятов, необходимый для функционирования цикла Кальвина, осуществляется по флоэме. При интенсивном фотосинтезе углеводы могут полимеризоваться, и при этом в хлоропластах формируются крахмальные зёрна. Газообмен (поступление углекислого газа и выделение кислорода) осуществляется путём диффузии через устьица (некоторая часть газов движется через кутикулу).
Фотосинтез на тканевом уровне
На тканевом уровне фотосинтез у высших растений обеспечивается специализированной тканью — хлоренхимой. Она располагается близ поверхности тела растения, где получает достаточно световой энергии. Обычно хлоренхима находится непосредственно под эпидермой. У растений, растущих в условиях повышенной инсоляции, между эпидермой и хлоренхимой может располагаться один или два слоя прозрачных клеток (гиподерма), обеспечивающих рассеивание света. У некоторых тенелюбивых растений хлоропластами богата и эпидерма (например, кислица). Часто хлоренхима мезофилла листа дифференцирована на палисадную (столбчатую) и губчатую, но может состоять и из однородных клеток. В случае дифференцировки наиболее богата хлоропластами палисадная хлоренхима.

Хлоропласты в клетках листа
Хлоропласты
Хлоропласты отделены от цитоплазмы двойной мембраной, обладающей избирательной проницаемостью. Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые, соединяясь друг с другом, образуют тилакоиды, которые, в свою очередь, группируются в стопки, называемые гранами. На мембранах тилакоидов располагаются молекулы хлорофилла и других вспомогательных пигментов (каротиноиды). Поэтому их называют фотосинтезирующими мембранами. Внутритилакоидное пространство отделено и не сообщается с остальной стромой; предполагается также, что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (с седиментацией типа 70 S), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения ): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённые состояния, при изменённом — триплетное первое и второе.
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла, в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
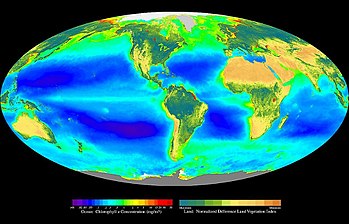
Сложноцветное изображение, показывающие глобальное распределение фотосинтеза, включая фитопланктон и наземную растительность
Фотосинтез составляет энергетическую основу всего живого на планете, кроме хемосинтезирующих бактерий.
Фотосинтез совершается в зеленых частях наземных растений и в водорослях. За один год зеленые водоросли выделяют в атмосферу Земли 3,6⋅1011 тонн кислорода, что составляет 90% всего кислорода, вырабатываемого в процессе фотосинтеза на Земле. Фотосинтез — самый массовый биохимический процесс на Земле [7] .
Возникновение на Земле более 3 млрд лет назад механизма расщепления молекулы воды квантами солнечного света с образованием O2 представляет собой важнейшее событие в биологической эволюции, сделавшее свет Солнца главным источником энергии биосферы.
Фототрофы обеспечивают конверсию и запасание энергии термоядерных реакций, протекающих на Солнце, в энергию органических молекул. Солнечная энергия при участии фототрофов конвертируется в энергию химических связей органических веществ. Существование гетеротрофных организмов возможно исключительно за счёт энергии, запасённой фототрофами в органических соединениях. При использовании энергии химических связей органических веществ гетеротрофы высвобождают её в процессах дыхания и брожения.
Фотосинтез является основой продуктивности как сельскохозяйственных растений, так и животной пищи.
Энергия, получаемая человечеством при сжигании биотоплива (дрова, пеллеты, биогаз, биодизель, этанол, метанол) и ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез служит главным входом неорганического углерода в биогеохимический цикл.
Большая часть свободного кислорода атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни существовать на суше.
Эффективность фотосинтеза — доля световой энергии, преобразуемая организмами в химическую в процессе фотосинтеза. Фотосинтез можно упрощённо описать с помощью химической реакции
Начиная с солнечного света, падающего на лист:
- 47 % энергии утрачивается, так как часть фотонов находится за пределами диапазона в 400—700 нм (если считать, что хлорофилл поглощает фотоны от 400 до 700 нм с эффективностью 100 %)
- 30 % теряется вследствие неполного поглощения фотонов хлоропластами, их отражения или поглощения другими компонентам клетки
- 24 % поглощенной энергии теряется из-за переноса энергии коротковолновых фотонов до уровня 700 нм
- 68 % используемой энергии теряется при превращении в D-глюкозу
- 35-45 % глюкозы потребляется листьями в процессах дыхания и фотодыхания.
Говоря другими словами: 100 % солнечного света → биодоступная радиация (400—700 нм) составляет 53 %, а 47 % оставшейся радиации не используется → 30 % фотонов теряются из-за неполной абсорбции 37 % (поглощенной энергии фотонов) → 24 % теряется в ходе переноса по антенным комплексам до уровня энергии 700 нм, оставляя 28,2 % энергии света, собранной хлорофиллом → 32 % преобразуются в АТФ и НАДФН, а затем в D-глюкозу, оставляя 9 % (сахар) → 35-40 % сахара потребляется листьями в процессе дыхания и фотодыхания, 5,4 % энергии идёт на чистый прирост биомассы.
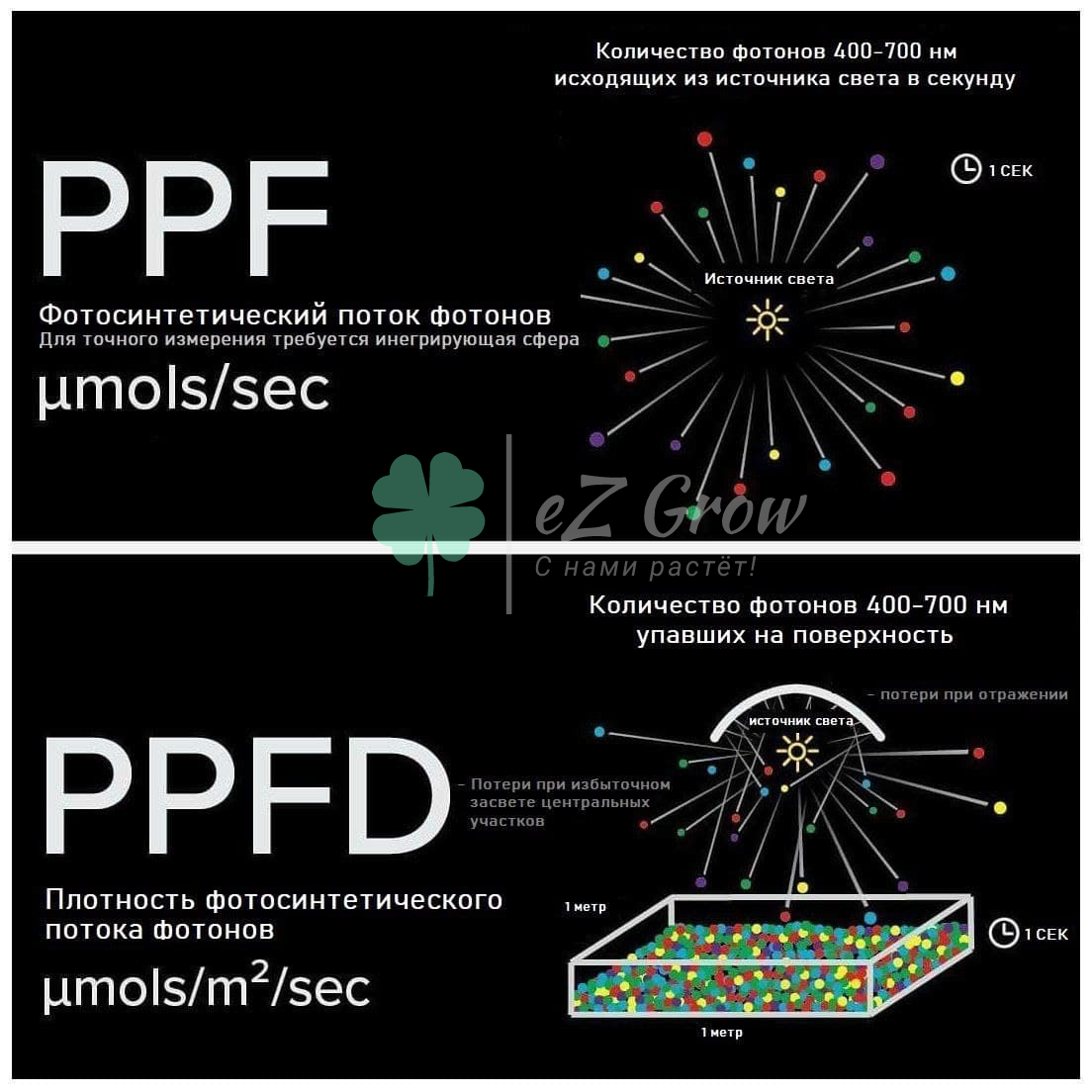
Многие растения тратят большую часть оставшейся энергии на рост корней. Большинство культурных растений запасают от ~0,25 % до 0,5 % энергии солнечного света в виде биомассы (кукурузные зерна, картофельный крахмал и др.). Исключением является только сахарный тростник, который способен запасать до 8 % солнечной энергии.
Интенсивность фотосинтеза линейно возрастает с увеличением интенсивности света, но постепенно достигает насыщения. В зависимости от культуры, при освещенности свыше 10000-40000 люкс (подразумевается солнечный свет) прекращается нарастание фотосинтеза. Таким образом, большинство растений может использовать только ~10-20% от общей энергии полуденного солнечного света. Однако дикие растения (в отличие от лабораторных образцов) имеют много избыточных, хаотически ориентированных листьев. Это даёт возможность держать среднюю освещенность каждого листа значительно ниже уровня полуденного пика освещённости, что позволяет растению достичь уровня ближе к ожидаемым результатам лабораторных испытаний, при относительно ограниченной освещённости.
Только если интенсивность света выше определённого значения, называемого световой точкой компенсации, растение усваивает больше углекислого газа, чем выделяет в результате клеточного дыхания.
Системы измерения фотосинтеза не способны непосредственно измерять количество света, поглощенное листом. Тем не менее, кривые отклика на свет, которые можно измерить и построить, позволяют сравнивать эффективность фотосинтеза у разных растений.
Фотосинтез

Фотоси́нтез, процесс преобразования зелёными растениями и некоторыми бактериями энергии света в энергию химических связей органических соединений:
C O 2 + H 2 A ⟹ h ν C H 2 O + 2 A , CO_2+H_2A\overset \nu>CH_2O+2A, C O 2 + H 2 A ⟹ h ν C H 2 O + 2 A , где H2A – восстановитель, который окисляется до A.
У высших растений, и цианобактерий восстановителем служит вода (H2O), а 2A – это кислород O2. В основе фотосинтеза лежат окислительно-восстановительные реакции , в ходе которых электроны переносятся от донора -восстановителя (H2O) к акцептору (CO2) с образованием восстановленных соединений ( углеводы ) и выделением молекулярного кислорода (O2). Такой фотосинтез называют оксигенным. Фотосинтезирующие бактерии, использующие в качестве донора-восстановителя не воду (H2O), а сероводород (H2S), аммиак (NH3), серу (S2), метан (CH4), молочную кислоту и другие легкоокисляемые соединения, O2 не выделяют.
Процесс фотосинтеза происходит в листьях , зелёных клетках стеблей , цветков , околоплодниках, черешках и даже в освещаемых корнях некоторых растений. Органеллой фотосинтеза является хлоропласт (у водорослей – хроматофор), который содержит различные пигменты – вещества, избирательно поглощающие свет в видимой части спектра .
Пигменты, участвующие в фотосинтезе высших растений, делятся на две группы: хлорофиллы (основные пигменты) и каротиноиды (вспомогательные пигменты). У пурпурных и зелёных бактерий, а также цианобактерий основными пигментами, участвующими в фотосинтезе, являются бактериохлорофиллы . У красных водорослей и цианобактерий роль вспомогательных ферментов выполняют фикобилины . У галобактерий фотосинтез осуществляется с помощью пигмент-белкового комплекса – бактериородопсина .
Развитие учения о фотосинтезе
В 1771 г. английский натуралист-любитель Дж. Пристли , проводя опыты с растениями, помещёнными в опрокинутый стеклянный кувшин, установил, что они способны «улучшать» воздух, «испорченный» при дыхании или горении свечи. В 1774 г. французским естествоиспытателем, основоположником современной химии А. Лавуазье , был получен молекулярный кислород.
В 1779 г. голландский врач Я. Ингенхауз показал, что зелёные растения выделяют O2 только на свету, однако при высоких температурах и интенсивности света они поглощают O2 и выделяют CO2.
В 1782 г. швейцарский естествоиспытатель Ж. Сенебье установил, что растения на свету не только выделяют O2, но и поглощают CO2. Процесс поглощения CO2 растениями Ж. Сенебье назвал «углеродным питанием». В 1800 г. им был издан трактат Physiologie végétale, в котором впервые предложен термин «физиология растений», а также определены задачи и методы новой науки.
В 1817 г. французские химики П. Пеллетье и Ж. Каванту выделили из листьев зелёное вещество и назвали его хлорофиллом. Важной ступенью в истории изучения фотосинтеза стало утверждение немецкого физика Р. Майера (1845) о том, что зелёные растения преобразуют энергию солнечного света в энергию химических соединений.
В 1865 г. немецкий учёный Ю. Сакс установил, что на свету в листьях образуется крахмал и он локализован в хлоропластах.
В 1871 г. российский естествоиспытатель К. А. Тимирязев , исследуя энергетику фотосинтеза, предположил, что хлорофилл способен к окислительно-восстановительному превращению. Позднее им была сформулирована идея о космической роли фотосинтеза как единственного процесса на Земле , благодаря которому солнечная энергия улавливается растением и трансформируется в энергию органических соединений (углеводов).
В 1881 г. немецкий физиолог Т. Энгельман экспериментально доказал, что кислород образуется в хлоропластах.
В 1893 г. российский биохимик А. Н. Бах предположил, что источником кислорода, образующегося при фотосинтезе, является вода.
Российский учёный М. С. Цвет в 1903 г. изобрёл новый метод разделения сложных веществ на компоненты – хроматографию , с помощью которой установил наличие двух модификаций хлорофилла: хлорофилл a и хлорофилл b, а также разделил жёлтые пигменты листа на три фракции.
В 1905 г. английский физиолог Ф. Блекман высказал предположение, что процесс фотосинтеза включает в себя световые реакции и не зависящие от света темновые реакции, для которых требуются вещества, образовавшиеся в световых реакциях.
Немецкий химик Р. Вильштеттер в 1906–1914 гг. определил элементарный состав хлорофиллов a и b. В 1915 г. за установление химического состава хлорофилла он был удостоен Нобелевской премии .
В 1914 г. один из основоположников отечественной физиологии растений, А. А. Рихтер , определил, что общая продолжительность акта фотосинтеза определяется темновой фазой. В 1920 г. немецкий биохимик О. Варбург доказал, что продолжительность световой фазы во много раз меньше темновой. В 1932 г. американскому учёному Р. Эмерсону удалось измерить продолжительность фаз фотосинтеза и показать, что скорость световых реакций во много раз выше темновых и они не зависят от температуры. Немецкий биохимик Х. Фишер в 1930–1940 гг. полностью расшифровал структурную формулу хлорофиллов a и b. В 1937 г. английский учёный Р. Хилл привёл биохимическое доказательство существования световой и темновой фаз фотосинтеза.
Прямые экспериментальные доказательства фотоокисления воды были получены советскими учёными А. П. Виноградовым и Р. В. Тейс в 1945 г. Они также показали, что при фотосинтезе кислород выделяется при разложении воды, а не углекислого газа.
В 1954 г. американский биохимик и физиолог растений Д. Арнон установил, что световая фаза фотосинтеза протекает в тилакоидах хлоропласта, а темновая – в строме . Он обнаружил, что освещение растворов хлоропластов в присутствии молекул аденозиндифосфата (АДФ) и фосфата приводит к образованию аденозинтрифосфата (АТФ). Этот процесс был назван фотосинтетическим фосфорилированием .
В 1956 г. группе американских учёных во главе с М. Калвином удалось раскрыть сущность темновых реакций фотосинтеза, за что им была присуждена Нобелевская премия (1961).
В 1957 г. Р. Эмерсон, изучая зависимость фотосинтеза от длины волны света, высказал идею о существовании двух фотосистем . В 1961–1966 гг. английский биохимик П. Митчелл предложил гипотезу, объясняющую сопряжение транспорта электронов с фосфорилированием, известную сейчас как «хемиосмотическая теория Митчелла», за которую он был удостоен Нобелевской премии (1978).
В 1966 г. австралийские исследователи М. Хетч и Ч. Слэк предложили законченную схему цикла темновых реакций у C₄-растений (C4-фотосинтез). В 1972 г. американский учёный Э. Толберт открыл фотодыхание .
Дальнейшее изучение процессов фотосинтеза показало, что свет активирует некоторые ферменты, катализирующие реакции темновой фазы. Кроме того, для многих ферментов темновой фазы была обнаружена зависимость от циркадных ритмов .
Существенные успехи, достигнутые мировой наукой за последние десятилетия, связаны с разработкой представлений о структурно-функциональной организации фотосинтетического аппарата, изучением молекулярных механизмов преобразования энергии, путей превращения углерода и др.
В настоящее время традиционное деление процесса фотосинтеза на две фазы – световую и темновую – часто заменяют на «первичные процессы фотосинтеза» и «восстановление CO2, NO3 — и SO4 2- » соответственно.
Первичные процессы фотосинтеза (световая фаза фотосинтеза)
Структура и функции фотосистем
У растений и цианобактерий световая фаза фотосинтеза включает в себя фотофизический и фотохимический процессы, которые требуют энергии света и осуществляются в мембранах обоих типов тилакоидов, где локализованы фотосинтетические системы – фотосистема I (ФС I) и фотосистема II (ФС II), а также компоненты электронн-транспортной цепи (ЭТЦ) и АТФазный комплекс.
Фотосистемы связаны между собой, но различаются по составу белков , пигментов и оптическим (спектральным) свойствам входящих в них пигментов. ФС I локализована в тилакоидах стромы и гран, а ФС II – в тилакоидах гран.
Каждая фотосистема представляет собой пигмент-белковый комплекс, куда входят реакционный центр (РЦ), антенный, или светособирающий, комплекс (ССК) и переносчики электронов. У высших растений выделено два реакционных центра, которые включают ассоциированные с белком молекулы пигмента-ловушки П700 (ФС I) или П680 (ФС II), представляющие собой димеры (т. е. две молекулы) хлорофилла a с максимумом поглощения света в наиболее длинноволновой части солнечного спектра – 700 или 680 нм соответственно.
В реакционном центре находится всего 1 % фотосинтетических пигментов, которые непосредственно участвуют в преобразовании энергии. Остальные 99 % пигментов играют вспомогательную роль, образуя ССК, в которых связаны с белками.
Два ССК расположены в реакционном центре ФС I и ФС II и связывают только молекулы ß- каротина хлорофилла a; два других – внешние ССК – связывают хлорофиллы a, b и каротиноиды. Каждый ССК содержит несколько сотен молекул хлорофиллов и каротиноидов.
В ССК молекулы хлорофиллов и каротиноидов выполняют функцию пигментов-сборщиков. Они поглощают энергию кванта света и передают её в реакционный центр пигментам-ловушкам П680 или П700. Передача (миграция) энергии осуществляется по принципу индуктивного резонанса благодаря очень тесному расположению пигментов. Установлено, что в ССК пигменты строго ориентированы друг относительно друга и расстояние между ними не превышает 1,15 нм. Скорость передачи энергии между пигментами составляет 10 -10 –10 -9 с. Миграция энергии происходит только в одном направлении, строго от каротиноидов к хлорофиллам b (эффективность передачи энергии составляет 40 %), от хлорофиллов b к хлорофиллам a (эффективность передачи энергии составляет 90 %) и, наконец, в РЦ к пигменту-ловушке П680 или П700. Таким образом, функция пигментов – сборщиков ССК состоит в поглощении энергии кванта света и её передаче реакционному центру, содержащему особые молекулы хлорофилла a – П700 или П680. На этом фотофизический процесс фотосинтеза заканчивается.
Поглотив энергию кванта света, электроны молекулы хлорофилла поднимаются на более высокий энергетический уровень, а сама молекула переходит в возбуждённое состояние. Приобретённый избыток энергии (т. н. энергия возбуждения) может быть перенесён на другую молекулу пигмента, растрачен в виде тепла, на испускание кванта света ( флуоресценция , фосфоресценция ) или использован в окислительно-восстановительной реакции (в фотохимическом процессе фотосинтеза).
В последнем случае в реакционном центре энергия возбуждения хлорофилла используется для первичного разделения зарядов , т. е. отщепления электронов от молекулы пигмента (она окисляется, становится заряженной положительно) и передачи их первичному акцептору, который восстанавливается (получает отрицательный заряд) и далее передаёт электроны переносчикам ЭТЦ. При оптимальном освещении разделение зарядов в РЦ происходит до 200 раз в секунду. Таким образом, в реакционном центре энергия возбуждённого состояния хлорофилла преобразуется в энергию разделённых зарядов, т. е. превращается в химическую энергию.
В ЭТЦ порядок расположения молекул-переносчиков зависит от их способности присоединять или отдавать электроны, т. е. величины окислительно-восстановительного потенциала (E’0).
Переносчиками электронов являются следующие белки: железосерные , или Fe-S-белки, ферредоксин , цитохромы b6 и f (образуют цитохромный b6f-комплекс), пластохиноны , флавиновые ферменты, пластоцианин , а также пиридиннуклеотиды ( никотинамидадениндинуклеотидфосфат , НАДФ + ).
Известны три пути транспорта электронов в ЭТЦ: нециклический, циклический и псевдоциклический.
Нециклический транспорт электронов
Нециклический транспорт электронов является основным в процессе фотосинтеза. В нём принимают участие две фотосистемы, которые осуществляют последовательный перенос электронов от ФС II к ФС I. В ФС II нециклический транспорт электронов отвечает за продукцию окислительного потенциала, необходимого для расщепления молекулы воды, в ФС I является сильным восстановителем и отвечает за образование богатого энергией кофермента – восстановленного НАДФН.
Диаграмма, на которой переносчики электронов расположены в соответствии со своим E’0, имеет форму буквы Z, поэтому получила название Z-схемы фотосинтеза.

Электрон-транспортная цепь хлоропластов (Z-схема фотосинтеза). Электрон-транспортная цепь хлоропластов (Z-схема фотосинтеза). Молекула П700 (E’0 = +0,43 В) ФС I, поглотив квант света, переходит в возбуждённое состояние (П700 * , E’0 = −0,8 В) и приобретает способность отдавать электроны первичному акцептору – мономерной форме хлорофилла a. Отдав электрон, молекула П700 окисляется и приобретает положительный заряд П700 + . Далее электрон переходит через ряд переносчиков ЭТЦ (Fe-S-белки, ферредоксин), расположенных в порядке возрастания E’0, на конечный акцептор – НАДФ + , восстанавливая его до НАДФН. Источником электрона, заполняющего свободное место в П700 + , является молекула П680 (E’0 = +1,18 В) ФС II. При поглощении энергии кванта света молекула П680 переходит в возбуждённое состояние (П680 * , E’0 = −0,8 В) и приобретает способность отдавать электроны акцептору – феофитину (хлорофилл, в котором отсутствует магний ). Феофитин выполняет роль промежуточного акцептора электрона между первичным донором П680 и первичным акцептором электрона, пластохиноном QA. Далее электрон, теряя энергию, переходит через ряд белков-переносчиков ЭТЦ к П700 + .
Таким образом, компоненты ЭТЦ соединяют между собой две фотосистемы. Цепь транспорта электронов включает следующие белки-переносчики: пластохиноны, цитохромный b6f-комплекс и пластоцианин. На участке транспорта электронов через цитохромный b6f-комплекс к П700 + возникает электрохимический градиент протонов за счёт переноса протонов из стромы через мембрану во внутритилакоидное пространство, который используется для образования АТФ из АДФ и неорганического фосфата (Фн).
Подробный механизм образования АТФ изложен в хемиосмотической теории (гипотезе), которую предложил П. Митчелл. Процесс преобразования энергии квантов света в энергию химических связей молекул АТФ был назван фотосинтетическим фосфорилированием ( фотофосфорилирование ), в отличие от окислительного фосфорилирования, наблюдаемого в митохондриях . В данном случае процесс образования АТФ из АДФ и неорганического фосфата (Фн) сопряжён с нециклическим транспортом электронов, поэтому получил название нециклического фотофосфорилирования.
Донором недостающего электрона для П680 + служат ионы марганца , образующие активный центр (Mn-кластер) водоокисляющего комплекса ФС II, в который входят также ионы хлора и кальция . В процессе последовательной передачи 4 электронов от водоокисляющего комплекса через промежуточный переносчик тирозин Z (остаток тирозина одного из белков ФС II) к П680 + происходит фотоокисление 2 молекул воды, которое сопровождается образованием 4 протонов (переносятся во внутритилакоидное пространство) и выделением в атмосферу молекулы кислорода:
2 M n 4 + + 2 H 2 O → 2 M n 2 + + 4 H + + 4 e ˉ + O 2 . 2Mn^ + 2H_2O → 2Mn^ + 4H^+ + 4ē + O_2. 2 M n 4 + + 2 H 2 O → 2 M n 2 + + 4 H + + 4 e ˉ + O 2 . Таким образом, при нециклическом транспорте электронов происходит перенос электронов от воды (E’0 = +0,81 В) к НАДФ + (E’0 = −0,32 В). В процессе участвуют две фотосистемы, которые образуют единую систему переноса электронов. Продуктами нециклического транспорта электронов являются АТФ и НАДФН. Суммарное уравнение нециклического фотофосфорилирования следующее:
2 НАД Ф + + 2 H 2 O + 2 АДФ + 2 H 2 P O 4 + 8 h υ → 2 НАДФН + 2 H + + 2 АТФ + O 2 . 2НАДФ^+ + 2H_2O + 2АДФ + 2H_2PO_4 + 8h \upsilon → 2НАДФН + 2H^+ + 2АТФ + O_2. 2 НАД Ф + + 2 H 2 O + 2 АДФ + 2 H 2 P O 4 + 8 h υ → 2 НАДФН + 2 H + + 2 АТФ + O 2 .
Циклический транспорт электронов с участием ФС I
Циклический транспорт электронов с участием ФС I является эволюционно самым древним. Он происходит тогда, когда соотношение между НАДФН и НАДФ+ достаточно велико.
Входящий в состав реакционного центра димер хлорофилла a П700 (E’0 = +0,43 В), поглотив квант света, переходит в возбуждённое состояние (П700 * , E’0 = −0,8 В). Отдавая электрон акцептору, молекула П700 окисляется и приобретает положительный заряд П700 + . Первичным акцептором электронов является мономерная форма хлорофилла a. От него электроны передаются по цепи переносчиков (Fe-S-белок, ферредоксин, пластохиноны, цитохромный b6f-комплекс, пластоцианин) с постепенной потерей энергии и возвращаются к П700 + , который в данном случае выступает одновременно и донором, и акцептором электронов.
Одновременно с электронами пластохиноны транспортируют через тилакоидную мембрану протоны, что создаёт электрохимический градиент протонов. Выделяемая в ходе окислительно-восстановительных реакций энергия частично используется на образование АТФ (в частности, на участке ЭТЦ между цитохромами b и f). В данном случае процесс образования АТФ за счёт энергии света, сопряжённый с циклическим транспортом электронов, получил название циклического фотофосфорилирования. При циклическом транспорте электронов АТФ представляет собой единственный продукт световой фазы фотосинтеза. Циклическое фотофосфорилирование является дополнительным источником АТФ для восстановления CO2 в темновой фазе процесса фотосинтеза. Его активность повышается при нахождении растений в условиях абиотического стресса.
Циклический транспорт электронов с участием ФС II
Циклический транспорт электронов с участием ФС II представляет собой перенос электронов от восстановленных пластохинонов к П680 + . Он является дополнительным источником АТФ и происходит, когда повреждён водоокисляющий комплекс (следовательно, кислород не выделяется) или наблюдается высокая интенсивность света. Предполагают, что этот процесс может протекать при закрытых устьицах , т. к. CO2 для него не нужен.
Псевдоциклический транспорт электронов
Процесс псевдоциклического транспорта электронов активируется в условиях дефицита НАДФ + и при наличии высокой интенсивности света. В нём участвуют две фотосистемы и белки-переносчики ЭТЦ. При псевдоциклическом транспорте электроны переносятся от H2O на O2 с образованием супероксидрадикала (O2 ·- ), одной из форм активированного кислорода, восстановление которого происходит в ФС I или ФС II, при этом синтезируется АТФ (псевдоциклическое фотофосфорилирование). Часто происходит так, что в ходе процесса единственным продуктом является АТФ, потому что потребление O2 компенсирует его образование при фотоокислении H2O.
Биохимическое значение первичных процессов фотосинтеза (световой фазы)
У высших растений первичные процессы фотосинтеза (световая фаза) обусловлены вызванным поглощённым светом транспортом электронов, в результате которого происходит фотоокисление воды (разрыв химических связей восстановителя) с выделением молекулярного кислорода и преобразование электромагнитной энергии в энергию химических связей АТФ и НАДФН – первичных стабильных продуктов, которые в дальнейшем используются при восстановлении CO2 и образовании конечных продуктов фотосинтеза.
Восстановление CO₂, NO₃ — и SO₄² — (темновая фаза фотосинтеза)
Темновая фаза фотосинтеза включает в себя комплекс биохимических реакций восстановления атмосферного CO2, поступающего через межклетники листа в хлоропласты, а также восстановления корнем NO3 — и SO4 2- – до органических соединений с использованием продуктов световой фазы. Известны различные пути восстановления CO2: C3-цикл (C3-фотосинтез, цикл Калвина , пентозофосфатный цикл ), C4-цикл ( C ₄ -фотосинтез , цикл Хетча – Слака – Карпилова), САМ-фотосинтез (от англ. Crassulacean Acid Metabolism, кислотный метаболизм толстянковых ), фотодыхание (C2-фотосинтез, гликолатный цикл).
C₃-цикл восстановления CO₂
C3-цикл восстановления CO2 является основным у растений и происходит в ходе последовательных ферментативных превращений – цикла Калвина (назван по имени описавшего его М. Калвина).

Растения, у которых функционирует только C3-цикл, получили название C3-растений. Ферменты, осуществляющие фиксацию CO2 и его восстановление до сахаров, расположены в строме хлоропласта. Цикл Кальвина. Цикл Кальвина.
В цикле Калвина выделяют четыре стадии:
1. Стадия карбоксилирования. На этой стадии происходит присоединение CO2 к 5-углеродному сахару – рибулозо-1,5-бифосфату (РуБФ). Реакцию карбоксилирования осуществляет фермент рибулозобифосфаткарбоксилаза – самый распространённый белок на Земле. Кроме карбоксилазной активности, этот фермент проявляет оксигеназную активность (присоединяет O2 в процессе фотодыхания). Поэтому полное название фермента – рибулозобифосфаткарбоксилазаоксигеназа (РуБисКО). Продукт реакции карбоксилирования – нестойкое 6-углеродное соединение, которое распадается на 2 молекулы 3-фосфоглицериновой кислоты (3-ФГК). Молекула 3-ФГК является первичным продуктом включения CO2 в органические вещества и содержит 3 атома углерода, поэтому этот способ фиксации CO2 называют C3-цикл.
2. Стадия восстановления ФГК. При участии АТФ и НАДФН – продуктов световой фазы, 3-ФГК восстанавливается до 2 молекул 3-фосфоглицеринового альдегида (3-ФГА). Молекулы 3-ФГА частично превращаются в изомер дигидроацетонфосфат (ДГАФ).
3. Стадия регенерации рибулозо-1,5-бифосфата (РуБФ). 3-ФГА взаимодействует с ДГАФ с образованием фруктозо-1,6-бифосфата, которая в ходе последующих реакций превращается в рибулозо-5-фосфат. При участии АТФ рибулозо-5-фосфат фосфорилируется до РуБФ – первичного акцептора CO2, что и обеспечивает непрерывную работу цикла. Если цикл проходит 3 раза, то во второй стадии образуется 6 молекул 3-ФГА, из которых 5 используются для регенерации 3 молекул РуБФ, а оставшаяся молекула выходит из цикла и включается в синтетические процессы.
4. Стадия синтеза углеводных продуктов фотосинтеза. При оптимальной интенсивности света триозы (3-ФГА, ДГАФ) используются в хлоропласте для образования первичного крахмала или транспортируются в цитозоль и участвуют в синтезе сахарозы , тогда как в условиях низкой освещённости участвуют в образовании органических кислот .
Среди первых продуктов фотосинтеза обнаружены аминокислоты . Из промежуточных продуктов цикла Калвина могут образовываться жиры и липиды . С учётом того что основным продуктом фотосинтеза являются углеводы (гексозы), цикл Калвина можно представить в виде следующего суммарного уравнения:
6 РуБФ + 6 C O 2 + 12 НАДФН + 18 АТФ + 12 H + → 6 РуБФ + C 6 H 12 O 6 + 12 НАДФ + + 18 АДФ + 6 H 2 O + 18 Ф н . 6РуБФ + 6CO_2 + 12НАДФН + 18АТФ + 12H^+ → 6РуБФ + C_6H_O_6 + 12НАДФ+ + 18АДФ + 6H_2O +18Ф_н. 6 РуБФ + 6 C O 2 + 12 НАДФН + 18 АТФ + 12 H + → 6 РуБФ + C 6 H 12 O 6 + 12 НАДФ + + 18 АДФ + 6 H 2 O + 18 Ф н .
Поскольку в этом цикле (стадия регенерации РуБФ) образуются пентозы (например, рибулозо-5-фосфат, ксилулозо-5-фосфат), то его также называют пентозофосфатный цикл , или восстановительный пентозофосфатный цикл.
C₄-цикл восстановления CO₂
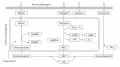
C4-фотосинтез (C4-цикл, C4-путь, цикл Хетча – Слака – Карпилова), комплекс биохимических Реакции темновой фазы фотосинтеза, протекающие у С₄-растений в клетках мезофилла. Реакции темновой фазы фотосинтеза, протекающие у С₄-растений в клетках мезофилла. реакций восстановления атмосферного углекислого газа (CO2), первыми продуктами которого являются органические кислоты ( яблочная , щавелевоуксусная и аспарагиновая ), содержащие 4 атома углерода.
Эволюционно C4-фотосинтез возник позже, чем C3-фотосинтез, поэтому у древесных растений он не обнаружен. Растения, имеющие C4-фотосинтез, называются C4-растениями.
C4-растения характеризуются высокой фотосинтетической продуктивностью и имеют преимущества перед C3-растениями при дефиците воды в почве .
CAM-фотосинтез

CAM-фотосинтез, двухэтапный комплекс биохимических реакций, которые разделены во Реакции темновой фазы фотосинтеза у САМ-растений. Реакции темновой фазы фотосинтеза у САМ-растений. времени: в тёмное время суток происходит связывание и запасание атмосферного углекислого газа (CO2) в вакуолях клеток (в виде яблочной кислоты), а днём – его высвобождение и включение в цикл Калвина.
CAM-метаболизм был впервые обнаружен у растений из семейства толстянковых (Crassulaceae). В настоящее время он описан у представителей 33 семейств. Растения, имеющие CAM-метаболизм, называют CAM-растениями.
Фотодыхание
Фотодыхание – активируемый светом процесс поглощения растениями O2 и выделения CO2. В присутствии высоких (около 2 %) концентраций O2 ключевой фермент C3-цикла РуБисКо функционирует как оксигеназа (присоединяет O2 к РуБФ).
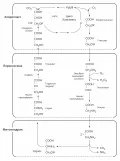
Фотодыхание. Фотодыхание. В ходе эволюции фотодыхание возникло как защита фотосинтетического аппарата растений от растущей концентрации атмосферного кислорода, выделяемого при фотосинтезе.
Фотодыхание осуществляется в результате взаимодействия трёх органелл – хлоропластов, пероксисом и митохондрий.
В процессе фотодыхания в хлоропластах и пероксисомах происходит поглощение O2, тогда как в митохондриях – выделение CO2.
Учитывая, что первичными продуктами фотодыхания являются фосфогликолат, гликолат и глиоксилат – двухуглеродные органические кислоты, этот процесс также называют C2-фотосинтезом, или гликолатным циклом. Фотодыхание является источником азотистых соединений – аминокислот глицина и серина .
Биологическое значение процесса фотосинтеза
Фотосинтез – единственный процесс в биосфере , ведущий к увеличению свободной энергии биосферы за счёт внешнего источника – Солнца и обеспечивающий существование как растений, так и всех гетеротрофных организмов, в том числе человека . Способность использовать солнечную энергию и вводить её в биосферные процессы и определяет космическую роль зелёных растений, о которой говорил и писал великий русский учёный К. А. Тимирязев.
Ежегодно в результате фотосинтеза на Земле образуется 160–200 млрд т органического вещества и выделяется 100–150 млрд т кислорода. Фотосинтетическое окисление воды является основным источником атмосферного кислорода. Кругооборот кислорода, углерода и других элементов, вовлекаемых в фотосинтез, создал и поддерживает современный состав атмосферы. Фотосинтез препятствует увеличению концентрации CO2 в атмосфере, предотвращая перегрев Земли вследствие парникового эффекта . Кислородно-озоновый экран атмосферы необходим для защиты от УФ-излучения.
Запасённая в продуктах фотосинтеза энергия (в виде различных видов топлива – торф , нефть , каменный уголь , древесина , природный газ ) является основным источником энергии для человека. Однако запасы ископаемого топлива ограниченны на планете. Предполагается, что в энергетике будущего фотосинтез может занять одно из первых мест в качестве неиссякаемого и не загрязняющего среду источника энергии (создание «плантаций» быстрорастущих растений с последующим использованием растительной массы для получения тепловой энергии или переработки в высококачественное топливо ).
Не менее важна роль фотосинтеза как основы получения продовольствия, кормов, технического сырья. Несмотря на высокую эффективность начальных стадий (около 95 %), в биомассу переходит менее 1–2 % солнечной энергии; потери обусловлены неполным поглощением света, лимитированием процесса на биохимическом и физиологическом уровнях.
Обеспечение растений водой, минеральным питанием, CO2, селекция сортов с высокой эффективностью фотосинтеза, создание благоприятной для светопоглощения структуры посевов и другие пути используются в целях реализации значительных резервов фотосинтетической продуктивности. Для ряда культур оправданны выращивание при полном или частичном искусственном освещении, биотехнологические способы получения растительной массы (особенно одноклеточных организмов ), аквакультура для некоторых водорослей и т. д. В связи с этим особенно актуальными становятся разработка теоретических основ управления фотосинтезом, исследование фотосинтеза как целостного процесса, закономерностей его регулирования и адаптации к внешним условиям.
В 21 в. перед учёными стоит глобальная задача – выяснить молекулярные механизмы преобразования энергии в процессе фотосинтеза, что необходимо не только для развития фундаментальной науки, но и для создания новых нанобиотехнологий . В качестве альтернативного источника энергии активно проектируются системы искусственного фотосинтеза (например, солнечные ячейки на основе компонентов фотосинтетического аппарата), проводится поиск высокоэффективных катализаторов, осуществляющих процесс окисления воды. Разработка искусственных фотохимических систем, способных генерировать молекулярный водород (т. н. фотоводород) и кислород за счёт электронов и протонов от воды и энергии солнечного света, обеспечит население Земли экологически чистым видом топлива и энергии. Достигнутые в мире в этом направлении успехи позволяют считать искусственный фотосинтез реальностью ближайшего будущего.
Опубликовано 9 ноября 2022 г. в 22:36 (GMT+3). Последнее обновление 9 ноября 2022 г. в 22:36 (GMT+3). Связаться с редакцией