Что такое поляризация клеток
Сортировка белка в поляризованных клетках эпителия
• В поляризованных клетках плазматическая мембрана состоит из отдельных доменов со специфическими наборами белков, и поэтому в этих клетках происходит дополнительная сортировка белков
• В зависимости от типа клеток, сортировка белков поверхности в поляризованных клетках может происходить в транс-Гольджи сети, эндосомах или в одном из доменов плазматической мембраны
• Процесс сортировки белков в поляризованных клетках происходит при участии специальных адаптерных комплексов, а также, вероятно, липидных рафтов (микродоменов) и лектинов
Плазматические мембраны большинства клеток многоклеточных организмов состоят из множества отдельных доменов, расположенных в непрерывном липидном бислое и различающихся между собой по биохимическим, структурным и функциональным характеристикам. Такие клетки считаются «поляризованными». Наиболее типичными примерами таких клеток являются эпителиальные клетки и нейроны.
Эпителиальные клетки выстилают все полости тела (например, кишечник, почки, воздухоносные пути) и, таким образом, обладают двумя поверхностями. Апикальный домен обращен в люмен органа, а базолатеральный граничит с кровью или с примыкающими клетками.
В эпителиальных клетках, специализированных на поглощении питательных веществ (например, клетки эпителия кишечника), на плазматической мембране апикального домена существуют небольшие выросты, которые называются микроворсинки. Они увеличивают общую поверхность клетки, что способствует увеличению поглощения ею питательных веществ.
Апикальная плазматическая мембрана в таких клетках обогащена мембранными белками, которые способствуют захвату клеткой таких питательных компонентов, как аминокислоты, сахара и другие молекулы. Также мембрана богата уникальными гликолипидами.
Напротив, базолатеральный домен содержит большинство мембранных белков (например, ЛНП-рецептор и ЭФР-рецептор) и липидов, находящихся в плазматической мембране неполяризованных клеток. Наряду с этим, переносчики, находящиеся в базолатеральном домене, транспортируют питательные компоненты из клетки в плазму крови.
Межклеточные контактные комплексы, состоящие из плотных контактов, адгезивных контактов и десмосом, разграничивают апикальные и базолатеральные домены. Одна из функций этих комплексов состоит в том, чтобы предотвратить возможность латеральной диффузии компонентов апикальной мембраны в базолатеральный домен, и наоборот.
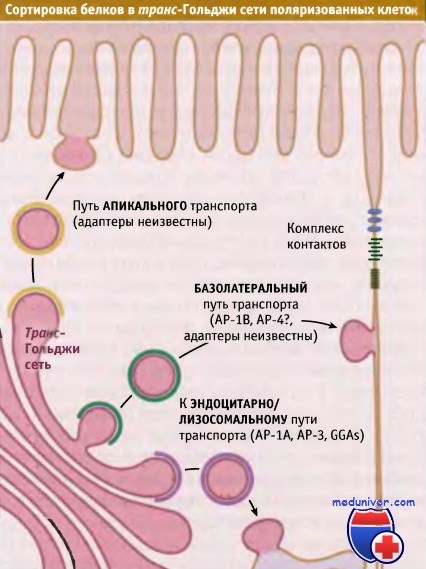
Три типа сортировки, происходящего в транс-Гольджи сети поляризованных клеток эпителия.
Белки сортируются для эндоцитоза и для транспорта через апикальную и базолатеральную мембраны.
Для каждого пути используются свои адаптерные комплексы.
Поэтому липиды и белки должны направляться в соответствующий домен. Транспорт по секреторному механизму и по пути эндоцитоза носит поляризованный характер. Это обеспечивает движение компонентов апикальной и базолатеральной мембран в соответствующих направлениях. Транспорт в базолатеральный домен происходит под действием адресных сигналов, отличающихся от сигналов для апикального транспорта.
В зависимости от того, предназначены вновь образованные мембранные белки для апикальной или базолатеральной поверхности, в транс-Гольджи сети они сортируются по разным типам везикул. Для многих типов поляризованных эпителиальных клеток процесс базолатерального транспорта исследован хорошо. В цитоплазматических доменах большинства базолатеральных белков присутствуют сигналы, содержащие остаток тирозина или два остатка лейцина.
Эти остатки узнаются адаптерными комплексами таким же образом, как это происходит для белков, которые транспортируются по механизму эндоцитоза или направляются в лизосомы. Последовательности, содержащие и иногда не содержащие остатки тирозина, узнаются в транс-Гольджи сети с участием АР-1В, специфичной для эпителиальных клеток изоформы клатринового адаптерного комплекса АР-1. АР-1В связывается с клатрином, образуя транспортную везикулу, ориентированную в базолатеральном направлении. Этот комплекс отличается от широко распространенного комплекса АР-1А только структурой μ-цепи. Субъединицы μ1А и μ1В обладают на 80% одинаковой структурой, однако связывают разные типы сигналов. Причина этих различий не установлена.
Цитоплазматические домены белков, предназначенных для апикальной плазматической мембраны, не содержат специфических сигналов. Вместо этого в домене люмена они содержат критические N- или О-сахарные остатки или имеют особый тип мембранного якорного домена, обеспечивающего их включение в апикально направляющиеся транспортные везикулы. Одним из примеров такого мембранного якоря является GPI, который добавляется к некоторым белкам в эндоплазматическом ретикулуме. Такие везикулы, транспортируемые в апикальном направлении, часто содержат уникальный липидный микродомен, называемый липидный рафт, который накапливает специфические белки, обеспечивая, таким образом, их селективное включение в апикальные везикулы.
Вероятно, этот же механизм в определенной степени позволяет разделять липиды, так что сложные гликолипиды, обнаруженные на апикальной, но не на базолатеральной поверхности эпителиальных клеток, также селективно включаются в эти везикулы.
Для некоторых мембранных белков, в которых отсутствует информация об адресе и которые транспортируются из транс-Гольджи сети и в апикальных, и базолатеральных везикулах, существует третий тип поляризованного адресования. Однако после попадания в плазматическую мембрану эти белки еще могут стать поляризованными. Это происходит в процессе домен-специфического удержания, при котором мембранный белок взаимодействует с предварительно поляризованным цитоскелетом.
Последний содержит белки, которые собираются на мембранных доменах в ответ, например, на такие сигналы, как прикрепление к другой клетке. Часто такой тип цитоскелета собирается в области межклеточных контактов, что обеспечивает асимметричность накопления мембранных белков. Мембранные белки, которые взаимодействуют с таким цитоскелетом, стабилизируются в соответствующем мембранном домене. Однако мембранные белки, попавшие в домен, который не обладает подходящим цитоскелетом, интернализируются путем эндоцитоза, деградируют в лизосомах, или рециклируют.
В последнем случае они получают еще одну возможность попасть в домен с соответствующим цитоскелетом.
Сортировка апикальных и базолатеральных белков также осуществляется в эндосомах. Поскольку эндоцитоз происходит как с апикальной, так и с базолатеральной поверхности эпителиальных клеток, интернализованные белки за счет рециклирования должны вернуться в те домены, из которых они вышли. Очевидно, ранние эндосомы могут вести себя как транс-Гольджи сеть, сортируя апикальные и базолатеральные белки в определенные везикулы рециклирования, которые направляются или к апикальной, или к базолатеральной мембране.
Фактически при сортировке по признаку поляризации в эндосомах используются те же сигналы, что и в транс-Гольджи сети. Чтобы при продолжающемся эндоцитозе эпителиальные клетки могли поддерживать полярность плазматической мембраны, необходимо поляризованное рециклирование содержимого эндосом. Фактически в эндосомах, и в частности, в рециклирующих эндосомах, может происходить биосинтетическая сортировка. Таким образом, в качестве промежуточных звеньев, на пути от транс-Гольджи сети к базолатеральной плазматической мембране, могут использоваться рециклирующие эндосомы. Это, вероятно, объясняет, почему оба пути используют один и тот же набор сигналов.
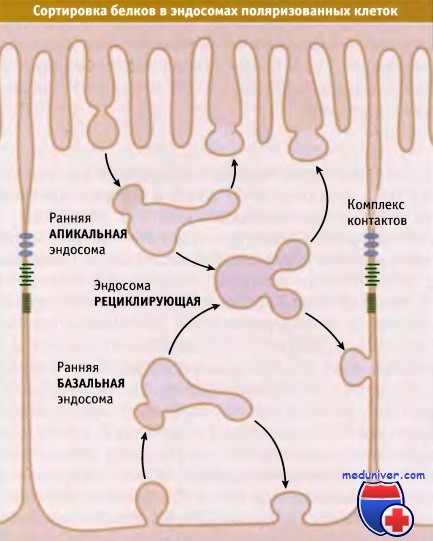
Молекулы, интернализованные из апикальной или базолатеральной мембран,
могут вернуться обратно в прежний домен мембраны прямым путем с участием ранних эндосом или направиться в другой домен при участии рециклирующих эндосом.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Локализация и функции адаптерных комплексов
- Судьба рецептора эндосомы после захвата клеткой
- Механизм созревания эндосом и их превращения в лизосомы
- Транспорт и сортировка лизосомальных ферментов в транс-Гольджи
- Сортировка белка в поляризованных клетках эпителия
- Образование секреторных гранул и их регуляция
- Перспективы изучения транспорта белков в клетке
- Строение ядра клетки и его функции
- Факторы влияющие на размер и форму ядра клетки
- Локализация хромосом в ядре клетки
Что такое поляризация клеток
Физики сумели измерить электрический потенциал, существующий между двумя сторонами клеточной мембраны. Он оказался равным 0,07 вольт. Можно сказать также, что этот потенциал равен 70 милливольтам, так как милливольт равен одной тысячной вольта. Конечно, это очень маленький потенциал по сравнению со 120 вольтами (120 000 милливольт) напряжения в сети переменного тока или по сравнению с тысячами вольт напряжения в линиях электропередачи. Но это все же удивительный потенциал, учитывая материалы, которые имеет в своем распоряжении клетка для построения электрических систем.
Любая причина, прерывающая деятельность натриевого насоса, приведет к резкому выравниванию концентраций ионов натрия и калия по обе стороны мембраны. Это, в свою очередь, автоматически приведет к выравниванию зарядов. Таким образом, мембрана станет деполяризованной. Конечно, это происходит при повреждении или гибели клетки. Но существуют, правда, три вида стимулов, которые могут вызвать деполяризацию, не причиняя клетке никакого вреда (если, конечно, эти стимулы не слишком сильны). К таким лам относятся механические, химические и электрические.
Давление — это пример механического стимула. Давление на участок мембраны приводит к а расширению и (по пока не попятным причинам) вызовет в этом месте деполяризацию. Высокая температура приводит к расширению мембраны, холод сокращает ее, и эти механические изменения тоже вызывают деполяризацию.
К такому же результату приводит воздействие на мембрану некоторых химических соединений и воздействие на нее слабых электрических токов. (В последнем случае причина деполяризации представляется наиболее очевидной. В конце концов, почему электрический феномен поляризации нельзя изменить с помощью приложенного извне электрического потенциала?)
Произошедшая в одном месте мембраны деполяризация служит стимулом для распространения деполяризации по мембране. Ион натрия, хлынувший в клетку в месте, где произошла деполяризация прекратилось действие натриевого насоса, вытесняет наружу ион калия. Ионы натрия меньше размерами и более подвижны, чем ионы калия. Поэтому в клетку входит больше ионов натрия, чем выходит из нее ионов калия. В результате кривая деполяризации пересекает нулевую отметку и поднимается выше. Клетка снова оказывается поляризованной, но с обратным знаком. На какой-то момент клеш приобретает внутренний положительный заряд, благодаря присутствию в ней избытка ионов натрия. На внешней стороне мембраны появляется маленький отрицательный заряд.
Противоположно направленная поляризация может служить электрическим стимулом, который парализует работу натриевого насоса в участках, примыкающих к месту первоначального стимула. Эти примыкающие участки поляризуются, потом происходит поляризация с обратным знаком и возникает деполяризация в более отдаленных участках. Таким образом, волна деполяризации прокатывается по всей мембране. В начальном участке поляризация с обратным знаком не может продолжаться долго. Ионы калия продолжают выходить из клетки, постепенно их поток уравнивается с потоком входящих ионов натрия. Положительный заряд внутри клетки исчезает. Это исчезновение обратного потенциала в какой-то степени реактивирует натриевый насос в этом месте мембраны. Ионы натрия начинают выходить из клетки, и в нее начинают проникать ионы калия. Данный участок мембраны вступает в фазу реполяризации. Так как эти события происходят во всех участках деполяризации мембраны, то вслед за волной деполяризации по мембране прокатывается волна реполяризации.
Между моментами деполяризации и полной ре-поляризации мембраны не отвечают на обычные стимулы. Этот период времени называется рефракторным периодом. Он длится очень короткое время малую долю секунды. Волна деполяризации, прошедшая через определенный участок мембраны, делает этот участок невосприимчивым к возбуждению. Предыдущий стимул становится в каком-то смысле единичным и изолированным. Как именно мельчайшие изменения зарядов, участвующие в деполяризации, реализуют такой ответ, неизвестно, но факт остается фактом — ответ мембраны на стимул изолирован и единичен. Если мышцу стимулировать в одном месте небольшим электрическим разрядом, то мышца сократится. Но сократится не только тот участок, к которому было приложено электрическое раздражение; сократится все мышечное волокно. Волна деполяризации проходит по мышечному волокну со скоростью от 0,5 до 3 метров в секунду, в зависимости от длины волокна, и этой скорости достаточно, чтобы создалось впечатление, что мышца сокращается, как одно целое.
Этот феномен поляризации-деполяризации-реполяризации присущ всем клеткам, но в некоторых он выражен больше. В процессе эволюции появились клетки, которые извлекли выгоды из этого явления. Эта специализация может пойти в двух направлениях. Во-первых, и это происходит весьма редко, могут развиться органы, которые способны создавать высокие электрические потенциалы. При стимуляции деполяризация реализуется не мышечным сокращением или другим физиологическим ответом, а возникновением электрического тока. Это не пустая трата энергии. Если стимул -это нападение врага, то электрический разряд может ранить или убить его.
Существует семь видов рыб (некоторые из них костистые, некоторые относятся к отряду хрящевых, являясь родственниками акул), специализированных именно в этом направлении. Самый живописный представитель — это рыба, которую в народе называют «электрическим угрем», а в науке весьма символическим именем — Electrophorus electricus. Электрический угорь — обитатель пресных вод, и встречается в северной части Южной Америки — в Ориноко, Амазонке и ее притоках. Строго говоря, эта рыба не родственница угрям, ее назвали так за длинный хвост, который составляет четыре пятых тела этого животного, длина которого составляет от 6 до 9 футов. Все обычные органы этой рыбы умещаются в передней части туловища длиной около 15 — 16 дюймов.
Более половины длинного хвоста занято последовательностью блоков модифицированных мышц, которые образуют «электрический орган». Каждая из этих мышц производит потенциал, который не превышает потенциал обычной мышцы. Но тысячи и тысячи элементов этой «батареи» соединены таким образом, что их потенциалы складываются. Отдохнувший электрический угорь способен накопить потенциал порядка 600 — 700 вольт и разряжать его со скоростью 300 раз в секунду. При утомлении этот показатель снижается до 50 раз в секунду, но такой темп угорь может выдержать в течение длительного времени. Электрический удар достаточно силен для того, чтобы убить мелкое животное, которыми питается эта рыба, или чтобы нанести чувствительное поражение животному более крупному, которое по ошибке вдруг решит съесть электрического угря.
Электрический орган — это великолепное оружие. Возможно, к такому электрошоку с удовольствием прибегли бы и другие животные, но эта батарея занимает слишком много места. Представьте себе, как мало животных имели бы крепкие клыки и когти, если бы они занимали половину массы их тела.
Второй тип специализации, предусматривающий использование электрических явлений, протекающих па клеточной мембране, заключается не в усилении потенциала, а в увеличении скорости распространения волны деполяризации. Возникают клетки с удлиненными отростками, которые представляют собой почти исключительно мембранные образования. Главная функция этих клеток — очень быстрая передача стимула от одной части тела к другой. Именно из таких клеток состоят нервы — те самые нервы, с рассмотрения которых началась эта глава.
Источник: Мир Энциклопедий Аванта+
Авторское право на материал
Копирование материалов допускается только с указанием активной ссылки на статью!
Похожие статьи
- Этапы синаптичнои передачи нервного импульса
- Характеристика спинно-мозговой (цереброспинальной) жидкости (СМЖ)
- ЭЛЕКТРОФИЗИОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ НЕЙРОГЛИИ
- Фоновая активность нервных клеток
- Клеточная мембрана
Информация
Посетители, находящиеся в группе Гости, не могут оставлять комментарии к данной публикации.
Краткие теоретические сведения
Биологическим объектам присущи пассивные электрические свойства: сопротивление и емкость. Биологические ткани и клетки обладают свойствами как проводников, так и диэлектриков. Проводимость этих объектов обусловливается наличием свободных ионов. Диэлектрические свойства биологических объектов и величина диэлектрической проницаемости определяются структурными компонентами и явлениями поляризации. Общей особенностью биообъектов является несоответствие между большим содержанием электролитов в клетках живых организмов и низкими величинами их электропроводности.
Экспериментально определяя сопротивление живых клеток и тканей, следует учитывать их чувствительность к действию электрического тока. При этом используют малые напряжения, чтобы не нарушать целостности образца.
Электропроводность клеток и тканей для постоянного тока
Электропроводность L – это величина, обратная сопротивлению R проводника:
где r – удельное сопротивление, l – длина проводника, S – сечение проводника.
Сопротивление R является коэффициентом пропорциональности между разностью потенциалов V и силой тока I:
При пропускании постоянного тока через живые ткани было установлено, что сила тока не остается постоянной во времени, хотя прикладываемое напряжение не изменяется. Сразу после подключения источника напряжения сила тока начинает непрерывно падать и устанавливается на уровне много ниже исходного (рис. 5.10).

Рис. 5.10. Изменение силы тока во времени при наложении на ткань постоянной разности потенциалов: а) отсутствие поляризации, б) наличие поляризации [1].
Это происходит из-за возникновения в образце электродвижущей силы противоположного направления – ЭДС поляризации P(t). При отключении образца от источника напряжения в цепи некоторое время будет наблюдаться ток обратного направления, вызванный этой ЭДС.
Таким образом, закон Ома для такого случая записывают как:
Остановимся на объяснении явления поляризации биологической ткани чуть подробнее.
Т. к. любой биологический объект имеет сложную организацию, влияние электрического поля и реакция на него будут различными на разных структурных уровнях.
Рассмотрим процессы, происходящие в биологических тканях при приложении внешнего электрического поля.
1. Как известно, электрическое поле вызывает упорядоченное движение свободных зарядов, в нашем случае – ионов. Следовательно, возникает электрический ток, что приводит, согласно закону Джоуля-Ленца, к выделению тепла. Этот процесс дает вклад в активную составляющую импеданса.
2. Также электрическое поле действует на связанные электрические заряды, не имеющие возможности свободного перемещения и на заряды, перемещение которых возможно лишь в малой ограниченной области. Однако воздействие на эти обе группы зарядов приводит к значительному эффекту – перераспределению в пространстве электрических зарядов или, другими словами, поляризации биологической ткани.
Поляризация по своей природе делится на несколько видов:
1. Электронная поляризация — это смещение электронов на своих орбитах относительно положительно заряженных ядер, что приводит к возникновению индуцированных, наведенных диполей. Время возникновения электронной поляризации после мгновенного наложения поля, называемое временем релаксации, равняется 10 -16 -10 -14 с. Возникающий диполь имеет небольшую величину.
2. Ионная поляризация – это смещение иона относительно кристаллической решетки, что также приводит к возникновению дипольного момента. Характерное время релаксации — 10 -14 -10 -12 с.
3. Дипольная (ориентационная) поляризация появляется вследствие ориентации полярных молекул в составе вещества в соответствии с направлением электрического поля. Имеет большое значение для таких полярных молекул как вода, спирты, белковые макромолекулы с диссоциированными группами и т.п. Время релаксации для данного типа поляризации совпадает со временем поворота молекул, а потому зависит от вязкости среды, температуры и радиуса молекул. В зависимости от указанных факторов это время изменяется в пределах 10 -13 -10 -7 с.
4. Макроструктурная поляризация возникает вследствие неоднородности электрических свойств вещества. При наличии слоев с различной электропроводностью после наложения электрического поля заряды двигаются до границы проводящего слоя и там накапливаются. В результате этого процесса проводящая часть ткани приобретает дипольный момент и ведет себя подобно гигантской поляризационной молекуле. Время релаксации этого типа — 10 -8 -10 -3 с.
5. Поверхностная поляризация происходит на поверхностях, имеющих двойной электрический слой. При наложении поля происходит перераспределение ионов диффузионной части двойного электрического слоя: частицы дисперсной фазы смещаются в одну сторону, а ионы диффузионного слоя – в другую, таким образом формируя наведенные диполи. Время релаксации поверхностной поляризации от 10 -16 до 10 -14 с.
6. Электролитическая поляризация возникает между электродами, опущенными в раствор электролита, при пропускании через них электрического тока. При наложении разности потенциалов на электроды происходит перераспределение потенциалопределяющих ионов в диффузионной части двойного электрического слоя: в области катода концентрация ионов (катионов) увеличится, а в области анода – уменьшится. В процессе дальнейшего изменения концентрации ионов в приэлектродных слоях могут начаться электрохимические реакции на электродах: ионы будут переходить из раствора на один электрод, а с другого электрода уходить в раствор. Таким образом, и в случае электролитической поляризации появление ЭДС поляризации обусловлено смещением зарядов, которое в данном случае проявляется как изменение концентрации ионов в приэлектродной зоне. Время релаксации данного типа поляризации измеряется величинами порядка 10 -4 -10 2 с.
Различают также несколько структурных уровней поляризации:
1. В макроскопических областях, ограниченных средами с низкой электропроводностью, под действием электрического поля происходит перемещение ионов межклеточной жидкости до границ области. Возникает большой дипольный момент, величина которого определяется размерами области и суммарным зарядом перемещенных ионов.
2. В пределах отдельной клетки происходит смещение ионов цитозоли до границ клеточной мембраны, имеющей низкую электропроводность. Дипольный момент отдельной клетки невелик (), но при значительной плотности клеток их суммарный дипольный момент сравним по величине с дипольным моментом ткани.
3. Для ядер клеток и других внутриклеточных структур, имеющих собственную мембрану, механизм поляризации аналогичен клеточному. Дипольный момент мал по сравнению с клеточным.
4. Ориентационная поляризация белков и других макромолекул. Величина поляризации зависит от состояния этих молекул (свободные или связанные).
5. Ориентационная поляризация воды и электролитов. Величина поляризации наименьшая из всех.
Схематически разные структурные уровни поляризации представлены на рис. 5.11.

Рис. 5.11. Структурные уровни поляризации биологической ткани
Помимо прямого исследования зависимости поляризации от частоты, существует интегральный способ оценки этой зависимости – определение изменения поляризации во времени в ответ на ступенчатое (мгновенное) включение внешнего электрического поля. Если в момент времени t = 0 напряженность внешнего электрического поля возрастает от нуля до значения , то зависимость от времени поляризации имеет одинаковый характер для всех механизмов:
где – наибольшее значение поляризации и – время релаксации.
Таблица 5.2. Значения времени релаксации для разных типов поляризации , с.
| Уровень организации | Ткани, суспензии клеток | Клетки | Ядра и внутрикле-точные органеллы | Протеины и макромолекулы | Вода и электролиты |
| Время релаксации, с | 10 -1 –10 –3 | 10 -2 –10 -4 | 10 –5 –10 -7 | 10 -6 –10 -8 | 10 -11 |
В постоянном электрическом поле проявляются все типы поляризации и величина диэлектрической проницаемости максимальна.
Электропроводность клеток и тканей для переменного тока
Как известно, переменный ток – это ток, который изменяется как по величине, так и по направлению. Далее мы будем рассматривать только такие токи, которые подчиняются гармоническому закону:
где – мгновенное значение силы тока в момент времени ; – амплитуда тока; – циклическая частота колебаний.
При прохождении через ткани переменного тока, изменяющегося по гармоническому закону, падение напряжения на биологической ткани также изменяется по закону
где амплитуда напряжения и фазовый сдвиг между напряжением и током определяются параметрами биологической ткани.
В общем случае амплитудные значения силы тока и напряжения в цепи переменного тока связаны как
И если переписать предыдущее уравнение в виде:
то получим, что амплитудное значение тока определяется амплитудой напряжения и так на зываемым полным сопротивлением цепи или импедансом :
Величина называется реактивным сопротивлением. Причем, как известно, реактивное сопротивление есть разность индуктивного и емкостного сопротивлений цепи:
Однако, принято считать, что индуктивность биологических тканей пренебрежимо мала () а значит
Сдвиг фаз между током и напряжением можно вычислить из выражения:
Т. о. напряжение отстает по фазе от тока (), а величина фазового сдвига определяется отношением реактивной и активной составляющих импеданса
Значения угла сдвига фаз, полученные при частоте 1 кГц для различных биологических объектов приведены в табл. 5.3.
Таблица 5.3 – Угол сдвига фаз между током и напряжением для различных биологичеких объектов
| Объект | , град. |
| Кожа человека, лягушки | -55 |
| Нерв лягушки | -64 |
| Мышцы кролика | -65 |
Итак, для биологического объекта импеданс носит составной (комплексный) характер
Его активная составляющая связана, в первую очередь, с проводимостью внутренних жидких сред, являющихся электролитами. Различные процессы в тканях, сопровождающиеся необратимыми потерями энергии, также дают вклад в величину активной составляющей импеданса. Реактивная компонента определяется емкостными свойствами исследуемой ткани, в частности, емкостью биологических мембран. Кроме того, в емкостную составляющую импеданса дает вклад и область контакта стимулирующих электродов с биологическими тканями.
Как известно, емкость является коэффициентом пропорциональности между зарядом и потенциалом и определяется как отношение изменения заряда проводника Dq к изменению его потенциала Dj:
Для плоского конденсатора она определяется по формуле:
где S – площадь пластин, d – расстояние между ними.
Измеряемая емкость биологического объекта определяется поляризационной емкостью Cp, которая возникает в момент прохождения тока. Поляризационная емкость будет определяться следующим уравнением:
где в числителе задается изменение во времени заряда Dq, а в знаменателе – изменение потенциала Dj (I0 и It – начальное и конечное значение силы тока, I – ток мгновенный, R – сопротивление объекта).
К поляризационной емкости биологического объекта добавляется значительная по величине статическая емкость мембран (1 мкФ/см 2 ). Согласно последнему уравнению, величина поляризационной емкости зависит от времени действия поля, а значит на низких частотах может превышать величину статической емкости. На более высоких частотах (порядка 10 кГц) статическая емкость на несколько порядков выше поляризационной. А так как эти емкости соединены последовательно, то на высоких частотах общая величина емкости определяется меньшей по величине поляризационной емкостью.
Абсолютная величина (модуль) электрического импеданса определяется выражением:
При этом как абсолютная величина импеданса так и фазовый сдвиг являются функциями частоты переменного тока.
Зависимость электрического импеданса от частоты носит название дисперсии импеданса.
Наличие активных и реактивных свойств импеданса можно моделировать, используя эквивалентные электрические схемы. Рассмотрим некоторые из них (рис. 5.12).
Для схемы с последовательным соединением активного и реактивного сопротивлений (рис. 5.12, а), значение импеданса может быть найдено с помощью выражения:
Однако в области низких частот эта схема имеет существенное расхождение с экспериментальными данными: при , что явно носит нефизический характер.
При параллельном соединении активного и реактивного сопротивлений (схема на рис. 5.12, б) полное сопротивление участка цепи задается выражением:
Полученное математическое выражение удовлетворительно описывает частотную зависимость величины импеданса на низких частотах, но при , что не соответствует опытным данным
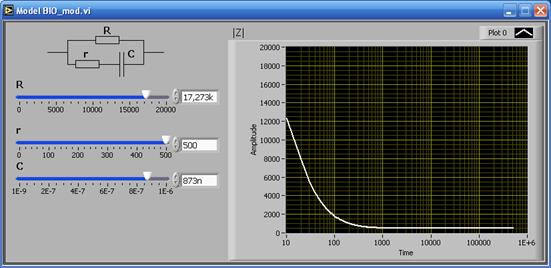
Наилучшее согласие с экспериментом обеспечивает эквивалентная схема, приведенная на рис. 5.12, в: на низких частотах величина импеданса определяется сопротивлением , на высоких частотах – параллельным соединением сопротивлений и . Можно показать, что в этом случае
Тогда видно, что в области низких частот, т. е. при , а в области высоких частот при
   |
а |
| б | |
| Рис. 5.13. Частотная зависимость импеданса : а – для эквивалентной схемы при различном соотношении активной и реактивной составляющих; б – экспериментальная зависимость для биологической ткани |
Как уже говорилось, и и (а следовательно и ), обладают свойством дисперсии. Вид кривой дисперсии импеданса (рис. 5.13, б) зависит от структурной целостности и от уровня обменных процессов.
Рассмотрим для начала емкостные свойства. Величина емкостного (реактивного) сопротивления конденсатора описывается формулой , а сама величина емкости для плоского конденсатора – .
Характеристикой реакции диэлектрической среды на электрическое поле является относительная диэлектрическая проницаемость – эта величина, определяющая, во сколько раз напряженность электростатического поля в данной точке среды меньше той напряженности электрического поля, которая возникла бы в вакууме при той же суперпозиции зарядов, создающих поле.
Вектором поляризации называется суммарный дипольный момент единицы объема среды :
В изотропной среде вектор поляризации параллелен вектору напряженности электрического поля и связан соотношением[С1]:
Отсюда легко видеть, что диэлектрическая проницаемость связана с поляризацией и напряженностью поля E соотношением:
Таким образом, именно зависимость поляризации среды от частоты может объяснить особенности поведения дисперсионной кривой реальной биологической ткани.
При увеличении частоты переменного тока (электрического поля) промежуток времени действия электрического поля (≈1/2 периода) уменьшается. Если этот промежуток времени меньше времени релаксации какого-то типа поляризации, то данный тип поляризации дает малый вклад в общее значение поляризации или совсем не проявляется. Этим объясняется зависимость диэлектрической проницаемости биологической ткани и, соответственно, импеданса от частоты.
Для оценки жизнеспособности ткани служит т. н. коэффициент поляризации:
где – значение импеданса на низкой частоте (обычно около 10 2 Гц); – на высокой частоте (>10 6 Гц).
Жизнеспособная ткань имеет , причем тем больше, чем выше уровень обменных процессов и чем лучше сохранена структурная целостность данной ткани. При отмирании ткани ее стремится к 1.
Итак, при прохождении переменного тока различной частоты через живую ткань отмечается явление дисперсии электропроводности, когда при повышении частоты тока сопротивление живой ткани уменьшается до некоторой предельной величины. Это явление обусловлено гетерогенностью клеточных структур и веществ цитоплазмы, а также структурированной организацией клетки. При действии внешнего электрического поля происходит ориентация всех заряженных частиц, молекул, ионов и диполей, а также структур с индуцированным зарядом против поля. Ионы при перемещении по полю накапливаются на внутренних и клеточной мембранах. Все эти факторы создают реактивное (в нашем случае емкостное) сопротивление внешнему электрическому току.
При изменении частоты тока происходит перезарядка мембран, переориентация молекул (т. е. поляризация). Поскольку у молекул и структур различные размеры и величины зарядов, то они имеют различное время релаксации . Поэтому с увеличением частоты тока часть из них (в первую очередь крупные частицы) не успевает переориентироваться и не участвует в создании внутренней ЭДС, вследствие чего ее величина уменьшается. В первом приближении эта теория удовлетворительно объясняет явление дисперсии электропроводности живой ткани, однако она не вскрывает причин изменения ее электропроводности при различных физиологических состояниях.
Виды поляризации.
Вещества обладают свободными и связанными зарядами. Свободные заряды – электроны и ионы под действием поля перемещаются от одного электрода к другому, создавая ток проводимости.
Особенности для биологических систем: в клетках свободные ионы могут перемещаться под действием поля в ограниченном объёме от одной мембраны до другой.
Связанные заряды под действием поля перемещаются только в очень ограниченных пределах, создавая при этом токи смещения.
Процесс перемещения связанных зарядов под действием электрического поля и образование ЭДС, направленной против внешнего поля, называется поляризацией.
Виды поляризации:
3) Дипольная (ориентационная);
1) Электронная поляризация представляет собой смещение электронов на своих орбитах относительно положительно заряженных ядер в атомах и ионах. В результате – атом или ион превращается в индуцированный, наведённый диполь с направлением, противоположным внешнему полю. Время возникновения электронной поляризации после мгновенного наложения поля, называется временем релаксации, равняется 10 -16 – 10 -14 с. Возникающий дипольный момент имеет небольшую величину.
2) Ионная поляризация – смещение иона относительно узлов кристаллической решётки. Вследствие этого возникает дипольный момент с направлением, противоположным внешнему полю. Время релаксации ионной поляризации 10 -14 — 10 -12 с.
3) Дипольная (ориентационная) поляризация. Если вещество содержит полярные молекулы и эти молекулы свободны, то под действием внешнего поля они ориентируются в соответствии с этим полем.

Этот вид поляризации имеет большое значение в веществах, молекулы которых обладают большим дипольным моментом (вода, спирт). Молекулы белков. а также других высокомолекулярных соединений, вследствие диссоциации ионогенных групп, а также вследствие адсорбции ионов обладают значительными дипольными моментами. Поэтому в растворах данных веществ структурная поляризация, обусловленная вращением их полярных молекул, имеет большое значение. Время возникновения дипольной поляризации – время релаксации – совпадает со временем поворота молекул. Время релаксации полярных молекул t зависит от вязкости среды m, T, радиуса молекулы r и вычисляется по формуле Стокса.

где К – постоянная Больцмана.
t дипольной поляризации изменяется в зависимости от указанных факторов в пределах от 10 -13 до 10 -7 с.
4) Макроструктурная поляризация возникает под действием электрического поля вследствие неоднородности электрических свойств вещества. Для её возникновения необходимо наличие слоёв с различной электропроводностью. Под действием поля свободные ионы и электроны, содержащиеся в проводящих субстанциях, перемещаются в пределах каждого включения до границы проводящего слоя. Дальнейшее перемещение свободных зарядов невозможно из-за низкой проводимости соседних слоёв. В результате этого включения проводящее включение приобретает дипольный момент и ведёт себя подобно гигантской поляризационной молекуле.

Время релаксации макроструктурной поляризации 10 -8 – 10 -3 с.
Биообъекты представляют собой гетерогенные структуры. Гетерогенность тканей обусловлена наличием мембран. К ним относятся клеточные мембраны и мембраны, окружающие клеточные органоиды и образующие эндоплазматическую сеть. Если цитоплазма обладает малым сопротивлением из-за наличия большого количества свободных ионов, то мембраны, наоборот, — большим (1000 Ом/см 2 ) в результате их малой проницаемости для ионов.
Макроструктурная поляризация происходит во всём объёме клеток, а не только на клеточной мембране, т.к. гетерогенность структуры имеется во всём объёме клеток. За счёт макроструктурной поляризации, которая играет основную роль в биологических объектах, диэлектрическая проницаемость тканей, измеренная в постоянном электрическом поле, достигает очень больших величин – нескольких миллионов.
5) Поверхностная поляризация – происходит на поверхностях, имеющих двойной электрический слой. При наложении внешнего поля происходит перераспределение ионов диффузионной части двойного электрического слоя: частицы дисперсной фазы смещаются в одну сторону, а ионы диффузионного слоя в другую. В результате частицы дисперсной фазы с противоионами диффузионного слоя превращаются в наведенные диполи. Время релаксации поверхностной поляризации лежит в пределах от 10 -3 до 1с.
6) Электролитическая поляризация возникает между электродами, опущенными в раствор электролита, при пропускании тока. До протекания тока концентрация потенциалопределяющих ионов у обоих электродов была одинакова, потенциалы электродов также были равны:
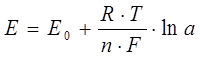
Е0 – нормальный потенциал электрода;
R – газовая постоянная;
n – валентность;
а – активность потенциалобразующих ионов при электродной зоне.
При наложении разности потенциалов на электроды произойдёт перераспределение потенциалобразующих ионов диффузионной части двойного электрического слоя: в области катода концентрация ионов (катионов) увеличится, а в области анода – уменьшится.
Обозначим активности ионов в приэлектродной зоне катода и анода через а1 и а2 . Тогда собственные потенциалы катода Ек и анода Еа будут:
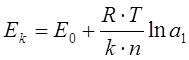
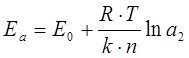
Между электродами возникает ЭДС поляризации , направленная против внешней ЭДС:

Таким образом, и при электролитической поляризации появление ЭДС поляризации обусловлено смещением зарядов, которое проявляется как изменение концентрации ионов в приэлектродной зоне. Время релаксации электролитической поляризации 10 -4 – 10 2 с.
Все описанные явления поляризации присущи биологическим объектам в той или иной мере. При наложении внешней разности потенциалов в тканях возникает противоположно направленное электрическое поле, которое значительно уменьшает внешнее поле и обусловливает высокое удельное сопротивление тканей постоянному току (10 6 — 10 7 Ом × см). Причем вначале возникают те виды поляризации, которые имеют меньшее время релаксации.
Удельное сопротивление тканей и жидкостей организма постоянному току (Ом×м)
1. Спинномозговая жидкость. 0,55
2. Кровь. 1,66
3. Мышцы. 2,00
4. Ткань мозговая и нервная. 14,3
5. Ткань жировая. 33,3
6. Кожа сухая. 10 5
7. Кость без надкостницы. 10 7
Все явления поляризации могут быть описаны с помощью диэлектрической проницаемости вещества.
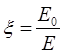
Е0 – напряжение электрического поля в вакууме;
Е — напряжение электрического поля в веществе.
3. Электропроводность клеток и тканей для переменного тока.
Биологические системы способны накапливать электрические заряды при прохождении через них тока, поэтому их электрические свойства описываются с помощью омического сопротивления и электрической ёмкости.

— электрическая ёмкость зависит от поляризации.
Для плоского конденсатора
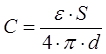
Ёмкость биологического объекта определяется поляризационной ёмкостью, которая возникает в момент прохождения тока. Поляризационная ёмкость показывает отношение изменения заряда объекта к изменению его потенциала при прохождении переменного тока.
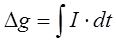

Тогда поляризационная ёмкость:
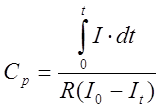
I – мгновенный ток;
I0, It – начальный и конечный ток.

К поляризационной ёмкости биологического объекта присоединяется значительная по величине статистическая ёмкость мембран (1мкФ/см 2 ). Как следует из уравнения (12) величина поляризационной ёмкости на низких частотах в зависимости от времени действия поля может значительно превышать величину статистической ёмкости. На более высоких частотах (10 кГц) статистическая ёмкость на несколько порядков выше поляризационной. А т.к. эти ёмкости соединены последовательно, то на высоких частотах общая величина ёмкости определяется меньшей по величине поляризационной ёмкостью.
Ткани организма состоят из клеток. При прохождении по ним электрического тока ионы перераспределяются. В области мембраны клеток возникает распределение зарядов, т.к. мембраны представляют собой препятствие для движения ионов. Это показано на эквивалентной схеме в виде С1 и С2. Резистор R1 отражает сопротивление цитоплазмы, а R2 сопротивление межклеточной жидкости.
Электрическая модель биологического объекта может быть представлена в виде различных эквивалентных схем (комбинации R и C).

Т.к. биологические объекты обладают как электрической проводимостью, так и ёмкостью, то они будут характеризоваться как активным, так и реактивным сопротивлением. Реактивное ёмкостное сопротивление:
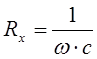
Суммарное сопротивление объектов называется импедансом.
Для последовательного соединения R и С импеданс определяется по формуле:
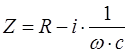

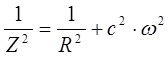
Z – импеданс;
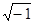
i – мнимая единица .
Т.к из формул следует, что импеданс объектов изменяется при изменении частоты тока – при увеличении частоты реактивная составляющая уменьшается.
Можно представить эквивалентную схему биологического объекта.
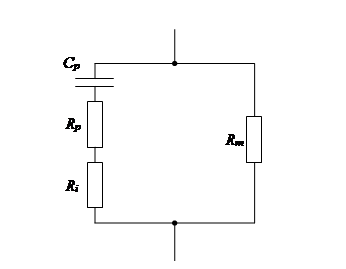
Поляризационные процессы на границе раздела – на мембране – обуславливают появление поляризацион-ного сопротивления Rp и ёмкости Ср зависят от результирующих ионных потоков через мембрану, т.е. от её проницаемости и статистической ёмкости.
Ri – сопротивление цитоплазмы;
Rm – сопротивление межклеточной жидкости.
В данной схеме Rp и Ср – частотнозависимые, а Ri и Rm – частотнонезависимые. На низких частотах общее сопротивление будет обусловлено в основном сопротивлением Rp, т.е. является в определённых условиях мерой проницаемости мембран. При увеличении частоты уменьшается Rp и Ср, что приводит к уменьшению эффекта поляризации границ раздела в биологических системах.
На высоких частотах (несколько МГц) Rp и Ср – станут стабильными и их можно исключить из схемы.
Частотнозависимый характер ёмкостного сопротивления является одной из причин зависимости импеданса биологических объектов от частоты тока, т.е. дисперсии импеданса.
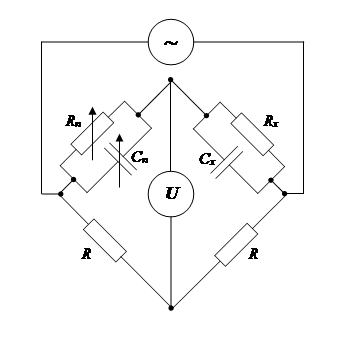
Электрические свойства тканей и клеток изучают с помощью мостовых схем.
R – омическое сопротивление уравновешенных плеч моста;
Rх и Cx – параметры объекта;
Сn и Rn – ёмкость и сопротивление, компенсирующие параметры объекта;
U – измерительный прибор.
При изучении частотных зависимостей сопротивления и ёмкости биологических объектов было обнаружено 3 области дисперсии.
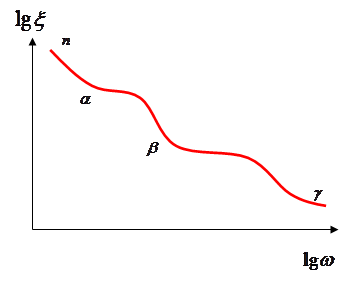
Частотная зависимость диэлектрической проницаемости мышечной ткани

С увеличением частоты уменьшается, образуя 3 зоны дисперсии a, b, и g.
a-дисперсия – область низких частот звукового диапазона до 1 кГц. Уменьшение диэлектрической проницаемости биологических систем обусловлено уменьшением поляризации поверхности клеток, т.к. сопротивление клеток для токов низкой частоты велико и высотное сопротивление будет представлено сопротивлениями электролитов Ri и RМ, т.е. при ВЧ сопротивление биологических объектов является показателем содержания в них свободных ионов.
b — дисперсия при частотах 10 3 – 10 7 Гц.
Наиболее полно электрические свойства биологических объектов в области b — дисперсии описываются теорией макроструктурной поляризации. Согласно этой теории, ёмкость и проводимость биологических объектов в данной области частот определяется гетерогенностью структуры – наличием мембран.
g — дисперсия на частоте >1000 МГц.
Уменьшение диэлектрической проницаемости в данном диапазоне обусловлено ослаблением эффекта поляризации, вызываемой диполями воды. g — дисперсия зависит от содержания свободной воды в биологических тканях. В области 400 МГц (между b и g дисперсиями) E тканей (кроме жировой, костной и мозговой) в пределах 40 – 60. При СВЧ >10 10 Гц эффект поляризации отсутствует. E будет небольшое значение, определяемое ионной и электронной поляризацией, имеющей малое время релаксации. На три основных дисперсионных явления, обусловленных поверхностной поляризацией, макроструктурной и дипольной поляризацией воды, накладываются сравнительно меньшие эффекты поляризации.
1) поляризация белковых и других органических молекул, обладающая дипольным моментами. Дисперсия на частотах в несколько МГц.
2) поляризация гидратных оболочек макромолекул (связанной воды). Частота (100 – 1000) МГц.
3) поляризация связанных групп макромолекул. Частота (100 – 1000) МГц.
Общая картина частотной зависимости электрических параметров сохраняется для всех тканей, что обусловлено единством структуры и химического состава клеток. Индивидуальные особенности (размеры и форма клеток, величина их проницаемости, содержание свободных ионов в клетках, содержание свободной воды) объектов обуславливают характер частотной зависимости.
4. Применение метода измерения электропроводности в биологии медицине.
Изучают проницаемость мембран методом электропроводности. Так, при воспалительных процессах в тканях на первых стадиях происходит увеличение сопротивления. Особенно сильно на низких частотах, когда сопротивление представлено в основном R межклеточных пространств. При воспалении происходит набухание клеток, объём межклеточных пространств уменьшается, а R – увеличивается. В первой стадии процесса структура клетки ещё не изменяется и ёмкость сохраняется прежней.
Следовательно, увеличение R при С = const всегда говорит о набухании тканей, а уменьшение R при С = const, наоборот – уменьшение объёма клетки.
В более поздних стадиях глубокие изменения структуры клетки, R и С уменьшаются. Таким образом, измерение электрических параметров тканей может средством для диагностики воспалительных процессов.
При повреждении тканей происходит увеличение проницаемости М, что приводит к падению R и С на низких частотах. На высоких частотах эффект отсутствует.
Таким образом, при повреждении и отмирании тканей дисперсия её электрических параметров снижается. При полной гибели – отсутствует.

2 – при нагревании до 50°С, 2 мин;
3 – то же в течении 4 мин;
4 – кипячение, 20 мин.
Тарусов для оценки жизнеспособности тканей предложил использовать коэффициент поляризации К.
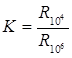
— отношение сопротивления на низких частотах к R на высоких.
К – характеризует величину дисперсии. Для печени млекопитающих К=9 – 10, для печени лягушки К=2 – 3, зависит от интенсивности обмена веществ. При отмирании ткани К уменьшается, а при гибели=1.
Метод регистрации изменений сопротивления органов, обусловленных изменением кровенаполнения, называется реографией. При наполнении органа кровью во время систолы сердца – R уменьшается, при диастоле – увеличивается. Можно изучать перераспределение крови между органами в различных условиях. Так исследуют кровообращение крови в почках, печени, сердце и др., кровоток в магистральных и более мелких сосудах.
Кровь обладает, практически, только активным сопротивлением, окружающие ткани обладают в значительной мере также и реактивным (ёмкостным) сопротивлением.
При определённой частоте переменного тока линии тока проходят через ткань будут концентрироваться в основном в кровеносных сосудах.
Изменение сопротивления участка ткани зависит от степени кровенаполнения и синхронно меняется с прохождением пульсовой волны.
График, изменяющийся со временем активной составляющей импеданса ΔR участка биоткани, называется реограммой.
Пусть ΔR – уменьшение сопротивления органа в момент прохождения по сосудам пульсовой волны, а R – его базовое сопротивление при отсутствии пульсовой волны.
Δv – изменение объёма органа за счёт притока крови (пульсовое приращение кровенаполнения), а V- базовый объём, то ΔR/R= ΔV/V
Таким образом пульсовое приращение ΔV=V · ΔR/R= a · ΔR
Где a = V/R – примерно постоянная величина во время диагностической процедура.
Сложность регистрации реограммы заключается в такой величине R» (0,05¸1)%R
Реограф состоит из мостовой схемы, в одно из плечей которого включен исследуемый орган пациента – RxCx. Конденсатор CP и резистор RP (магазин емкости и магазин сопротивления) предназначены для компенсации активных и реактивных свойств исследуемого органа пациента. R – сопротивление уравновешенных плечей моста. В реографе имеется генератор высокочастотных электрических колебаний 1, подающий напряжение на одну из диагоналей моста, усилитель 2 усиливающий изменение напряжения на другой диагонали моста, и регистрирующее 3 реограмму устройство. Регистрируется изменение напряжения в диагонали моста, пропорциональные измению сопротивления органа пациента ΔR. Применяется биполярная (двухэлектродная) и тетраполярная (четырехэлектродная) реография.

Реэнцефалография — метод исследования мозгового кровообращения. С помощью метода электропроводности можно изучать процессы связывания ионов молекулами белков. По уменьшению электропроводности можно вычислить количество ионов, связанных одной молекулой белка.